Protein folding at extreme temperatures: current issues
Texte intégral
Figure

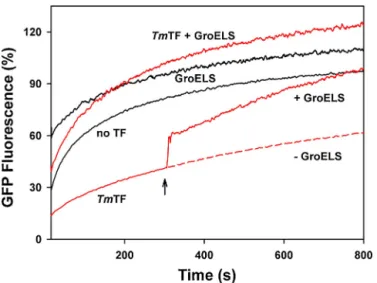



Documents relatifs
In another example, the acid-denatured states of ribonuclease, lysozyme and chymotrypsinogen, all three common model globular proteins for the study of protein folding, have shown
As previously mentioned, one of the main difficulties of the protein folding problem is the existence, in phase space, of a large number of local
For non chiral chains, a mean field study of this model shows that a classical 6 collapse transition occurs first; at lower temperature,.. nematic order
With a simple definition based on the well-known NKModel, the motivation is to make the IFP problem more accessible to optimisation specialists and model experts.Furthermore
S. Guillaumin, et al.. Muscle protein turnover in broiler chickens: effects of high ambient temperatures and dietary protein intake.. Muscle protein turnover in broiler..
While bounded model checking of timed automata has been explored in the literature based on the satisfiability of Boolean con- straint formulas over linear arithmetic
Finally, we examine how drugs targeting the protein folding activity of the ribosome could be active against mammalian prion and other protein aggregation-based diseases, making
In fact, whilst earlier comparison between the src and the spectrin SH3 domains suggested this class of proteins to fold via a robust two-state mechanism
