LA VOIE ASP/C5L2 DANS LE MÉTABOLISME
ÉNERGÉTIQUE
Thèse
Christian Roy
Doctorat en physiologie-endocrinologie
Philosophiae Doctor (Ph.D.)
Québec, Canada
© Christian Roy, 2014Résumé
L’obésité est un problème de santé majeur qui se traduit par un déséquilibre du métabolisme énergétique. La protéine stimulant l’acylation (ASP, C3adesArg) est une protéine produite par le tissu adipeux qui stimule la synthèse des triglycérides (TG) et le transport du glucose en se liant à son récepteur C5L2. Nos études précédentes ont établi que les souris avec le gène C3 invalidé (C3KO) sont des souris déficientes en ASP, hyperphagiques, mais de poids normal, affichant une dépense énergétique plus élevée que les souris sauvages. C’est à partir de ces résultats que nous avons basé notre objectif de déterminer le rôle de la voie ASP-C5L2 dans le métabolisme énergétique et cela autant au niveau du cerveau que des organes périphériques. Lors de notre première étude, nous avons observé qu’une injection centrale d’ASP au niveau du troisième ventricule chez des rats Wistar, amenait une diminution de la prise alimentaire et du poids corporel ainsi qu’une augmentation de l’expression d’ARNm du neuropeptide anorexigène pro-opiomélanocortine (POMC) dans la région du noyau arqué. Nos études révèlent également que les niveaux de lipides intramyocellulaires (IMCL) sont augmentés de six fois chez les souris déficientes en C5L2 (C5L2KO) par rapport aux souris sauvages (WT) suite à un régime riche en graisse. De plus, chez les sujets humains, l’expression protéique de C5L2 est réduite chez les patients diabétiques de type 2 par rapport aux témoins obèses. L'entraînement physique a augmenté l’expression protéique de C5L2 et la production d’ASP chez les hommes obèses insulino-résistants. Enfin, nos travaux avec les souris déficientes pour le récepteur du C5a (C5aRKO) montrent qu’après 12 semaines de diète riche en graisse et en sucre (DIO), les souris C5aRKO présentent une diminution du poids corporel ainsi que des dépôts adipeux gonadiques et inguinaux plus petits que leurs homologues WT. Des taux de triglycérides et d'acides gras non-estérifiés plasmatiques plus faibles et une clairance postprandiale plus rapide des triglycérides ont aussi été constatés. Nous avons donc démontré que la voie ASP/C5L2 représente une cible intéressante autant dans le contrôle de la prise alimentaire que dans le métabolisme des acides gras du muscle squelettique.
Abstract
Obesity is a major health problem that results in an imbalanced energy metabolism. Acylation Stimulating Protein (ASP, C3adesArg) is a protein produced by the adipose tissue, which stimulates triglycerides (TG) synthesis and glucose transport by binding to its receptor C5L2. Our previous studies have shown that mice with a disabled C3 gene (C3KO) are ASP deficient, hyperphagic but of normal weight and demonstrate greater energy expenditure than wild-type mice. From these results, our goal was to determine the role of the ASP-C5L2 pathway in energy metabolism. In our first study, we observed that a central injection of ASP in the third ventricle of Wistar rats brought a reduction in food intake and body weight gain and an increased mRNA expression of the anorexigenic neuropeptide POMC in the region of the arcuate nucleus. Our studies also showed that the levels of intramyocellular lipids (IMCL) were increased six-fold in C5L2-deficient mice (C5L2KO) compared to wild mice (WT) after a high-fat diet. Furthermore, in humans, the protein expression of C5L2 is reduced in patients with type 2 diabetes compared with obese controls. Physical training increased protein expression of C5L2 and ASP output in obese insulin-resistant men. Finally, our work with C5a receptor-deficient mice (C5aRKO) showed that after 12 weeks of a high-fat and high-sugar diet (DIO), the C5aRKO mice exhibited reduced body weight and smaller gonadal and inguinal fat mass than their WT counterparts. This was accompanied with lower plasma levels of non-esterified fatty acids and triglycerides and faster postprandial triglyceride clearance. Our studies have demonstrated that the ASP/C5L2 pathway represents an attractive target in the control of food intake and skeletal muscle fatty acid metabolism.
Table des matières
Résumé ... iii
Abstract ... v
Table des matières ... vii
Liste des tableaux ... xi
Liste des figures ... xiii
Liste des abréviations ... xv
Remerciements ... xxi
Avant-propos ... xxv
Articles présentés ... xxix
Introduction ... 1
1-Obésité ... 1
1.1-Définir le surpoids et l’obésité ... 1
1.2-Les causes de l’obésité et du surpoids ... 4
1.2.1 Dépense énergétique ... 6
1.2.1.1 Le métabolisme de repos ... 6
1.2.1.2 La thermogenèse adaptative ... 7
1.2.1.3 L’activité physique ... 8
1.2.2 Apport énergétique ... 9
1.3-Les pathologies reliées au surpoids et à l’obésité ... 10
1.3.1 Diabète de type 2 ... 11
1.3.2 Autres perturbations endocriniennes et métaboliques ... 13
1.3.3- Maladies cardiovasculaires ... 14
1.3.4- Problèmes respiratoires ... 14
1.3.5- Cancers ... 15
1.3.6- Stéato-hépatite non alcoolique ... 15
2-Tissus adipeux ... 16
2.1-Tissu adipeux brun ... 17
2.2-Tissu adipeux blanc ... 18
2.2.1 Emmagasinage de l’énergie ... 19 2.2.2 Organe endocrinien ... 23 2.2.2.1 Adiponectine ... 25 2.2.2.2 Leptine ... 28 2.2.2.3 La résistine ... 30 2.2.2.4 La retinol-binding protein-4 ... 32
2.2.2.5 Plasminogen activator inhibitor-1 (PAI-1) ... 33
2.2.2.6 Facteur de nécrose tumorale (TNF alpha) ... 34
2.2.2.7 L’interleukine 6 ... 35
2.2.3 Adipokine et le système nerveux central ... 36
2.2.4 Les adipokines, l’inflammation et le système immunitaire ... 41
3-Protéine stimulant l’acylation ... 43
3.1-L’origine ... 43
3.2-L’ASP et son récepteur ... 45
3.4-Rôle métabolique ... 49
3.4.1 Observations chez les humains ... 49
3.4.1.1- L’ASP et le tissu adipeux humain ... 51
3.4.2 L’ASP et le modèle animal ... 51
3.4.2.1 ASP et tissu adipeux animal ... 52
3.4.2.2 ASP et le muscle squelettique animal ... 53
Problématique ... 56
Objectifs des travaux ... 58
Objectifs spécifiques ... 58
Chapitre 1 ... 60
Acute injection of ASP in the third ventricle inhibits food intake and locomotor activity in rats ... 60 Résumé ... 61 Abstract ... 62 Introduction ... 64 Methods ... 66 Results ... 71
Discussion and Conclusion ... 74
Acknowledgements ... 77 Reference List ... 78 Tables ... 85 Figure legends ... 86 Figures ... 88 Chapitre 2 ... 92
Relationship of C5L2 receptor to skeletal muscle substrate utilization ... 92
Résumé ... 93
Abstract ... 94
Introduction ... 95
Materials and Methods ... 97
Results ... 101 Discussion ... 104 Acknowledgements ... 108 Ethical statement ... 108 References ... 109 Tables ... 115 Figure legends ... 118 Figures ... 120 Chapitre 3 ... 122
C5A receptor deficiency alters energy utilization and fat storage ... 122
Résumé ... 124
Abstract ... 125
Introduction ... 128
Materials and methods ... 131
Results ... 136
Discussion ... 141
Declaration of interest ... 145
Ethical statement ... 145 References ... 146 Figure legends ... 151 Tables ... 155 Figures ... 156 Supplementary tables ... 161
Discussion générale, perspectives de recherche et conclusion ... 162
Discussion générale ... 162
Perspectives de recherche ... 172
Conclusion ... 174
Liste des tableaux
Introduction
TABLEAU 1.CLASSIFICATION DU RISQUE POUR LA SANTÉ EN FONCTION DE L'INDICE DE MASSE CORPORELLE (IMC) ... 2
TABLEAU 2.RISQUES RELATIFS DE COMPLICATION MÉTABOLIQUE EN FONCTION DU TOUR DE TAILLE ... 4
TABLEAU 3-PROTÉINES ET FACTEURS NON-PROTÉIQUES SECRÉTÉS PAR LE TISSU ADIPEUX BLANC... 24
TABLEAU 4.AFFINITÉ DE L’ASP, DE C3A ET DE C5A POUR LES RÉCEPTEURS C5L2,C3AR ET C5AR. ... 47
Chapitre 1 TABLE 1.ICV INJECTION OF ASP DIMINISHES FOOD INTAKE AT EACH TIME INTERVAL. ... 85
TABLE 2.BLOOD PLASMA VALUES. ... 85
Chapitre 2 TABLE 1.MOUSE BODY WEIGHT AND FINAL PLASMA VALUES. ... 115
TABLE 2.MITOCHONDRIAL RESPIRATION RATES,ROS PRODUCTION AND OXPHOS CONTENT ... 116
TABLE 3.ANTHROPOMETRIC MEASUREMENTS AND PLASMA VALUES FOR OBESE AND TYPE 2 DIABETIC MEN ... 117
Chapitre 3 TABLE 1.BODY AND TISSUE WEIGHTS, FOOD INTAKE AND PLASMA PARAMETERS IN WILDTYPE AND C5AR KNOCKOUT MICE ... 155
SUPPLEMENTARY TABLE 1:SEQUENCES FOR PRIMERS USED IN RT-PCR.ALL SEQUENCES WERE OBTAINED THROUGH THE MOUSE PRIMER DEPOT RESOURCE SITE. ... 161
Liste des figures
INTRODUCTION
FIGURE 1.LA DISTRIBUTION RÉGIONALE DU TISSU ADIPEUX.. ... 3
FIGURE 2.LA BALANCE ÉNERGÉTIQUE.. ... 5
FIGURE 3.FACTEURS D’INFLUENCE SUR LE POIDS. ... 6
FIGURE 4.ILLUSTRATION DES MÉCANISMES LIANT L’OBÉSITÉ AU DIABÈTE DE TYPE 2. ... 12
FIGURE 5.ADIPOCYTE BLANC VS ADIPOCYTE BRUN. ... 16
FIGURE 6.DISTRIBUTION ANATOMIQUE DU TISSU ADIPEUX BLANC.. ... 19
FIGURE 7L’HYPOTHÈSE DU DÉBORDEMENT LIPIDIQUE.. ... 22
FIGURE 8.LE TISSU ADIPEUX, UN ORGANE ENDOCRINIEN... 23
FIGURE 9.LES CHANGEMENTS DANS LA SÉCRÉTION D’ADIPOKINES PROVOQUÉS PAR L'OBÉSITÉ CONDUISENT AU DÉVELOPPEMENT DE LA RÉSISTANCE À L'INSULINE, DU SYNDROME MÉTABOLIQUE, DU DIABÈTE DE TYPE 2 ET À DES TROUBLES CARDIOVASCULAIRES. ... 25
FIGURE 10.CENTRE DE CONTRÔLE DE LA PRISE ALIMENTAIRE AU NIVEAU DU SYSTÈME NERVEUX CENTRAL.… ... 38
FIGURE 11.SCHÉMATISATION DE LA PRODUCTION DE LA PROTÉINE STIMULANT L’ACYLATION (ASP). ... 44
FIGURE 12.SCHÉMA DU RÉCEPTEUR C5L2 HUMAIN.. ... 45
FIGURE 13.SCHÉMATISATION DE LA VOIE DE SIGNALISATION INTRACELLULAIRE PROPOSÉE POUR LA STIMULATION DE L’ASP SUR LA SYNTHÈSE DES TG. ... 49
Chapitre 1 FIGURE 1CENTRAL ASP INJECTION REDUCED FOOD INTAKE ... 88
FIGURE 2ICV INJECTION OF ASP HINDERS WEIGHT GAIN ... 88
FIGURE 3LOWER OXYGEN CONSUMPTION AND RQ LEVELS FOLLOWING AN ASP INJECTION ... 89
FIGURE 4CENTRAL INJECTION OF ASP ENHANCES SEDENTARY BEHAVIOR ... 90
FIGURE 5CENTRALLY INJECTED ASP IS NOT A MALAISE-INDUCING AGENT ... 90
FIGURE 6ASPICV INJECTION INFLUENCES NEUROPEPTIDE EXPRESSION IN THE ARCUATE NUCLEUS AS ASSESSED BY IN SITU HYBRIDIZATION ... 91
FIGURE 7RELATIVE EXPRESSION OF C5L2 IN THE BRAIN ... 91
Chapitre 2 FIGURE 1BODY WEIGHT AND IMCL QUANTIFICATION ... 120
FIGURE 2SKELETAL MUSCLE ENZYME ACTIVITY ... 120
FIGURE 3INSULIN SENSITIVITY ... 121
FIGURE 4SKELETAL MUSCLE C5L2 PROTEIN CONTENT IN HUMANS ... 121
Chapitre 3 FIGURE 1OXYGEN CONSUMPTION AND LOCOMOTOR ACTIVITY ... 156
FIGURE 2ALTERED BODY WEIGHT AND FAT PAD WEIGHT IN C5AR DEFICIENT MICE ... 157
FIGURE 3C5AR AND C5L2 EXPRESSION IN ADIPOSE, MUSCLE AND LIVER OF MICE ... 158
FIGURE 4METABOLIC RESPONSE TO GLUCOSE AND INSULIN TOLERANCE TESTS AND POSTPRANDIAL LIPID CLEARANCE IN C5ARKO MICE ... 158
FIGURE 5C3 AND C5A PLASMA LEVEL AND MRNA EXPRESSION IN MUSCLE AND LIVER OF GENES IMPLICATED IN SUBSTRATE UTILIZATION ... 159
Liste des abréviations
Abréviations Définitions
α-MSH Hormone stimulant le mélanocyte-alpha 3T3-L1 Souche de lignée cellulaire adipocytaire
acrp30 Adipocyte complement-related protein of 30 kDa
ACC Acétyl-COA carboxylase
ADKO Souris sans l’adipsine déficiente en ASP AdipoR1 Récepteur 1 de l’adiponectine
AdipoR2 Récepteur 2 de l’adiponectine
AG Acides gras
AGL Acides gras libres
AGNE Acide gras non estérifié AgRP Peptide relié à l’agouti
AMPc Adénosine 3',5'-monophosphate cyclique apM1 Adipose most abundant gene transcript1
AMPk Protéine kinase activée par l'adénosine monophosphate
apoB Apolipoprotéine B
ARC Noyau arqué
ARNm Acide ribonucléique messager
ASP Protéine stimulant l'acylation, C3adesArg
ATP Adénosine triphosphate
BKO Souris sans facteur B et déficiente en ASP
C3KO Souris sans le complément C3 donc déficientes en ASP C5aR Récepteur pour le C5a
C5aRKO Souris déficientes pour le récepteur du C5a C5L2 Récepteur C5aR like-2, GPR
C5L2KO Souris déficientes pour le récepteur C5L2
CART Neuromédiateur contrôlé par la cocaïne et l’amphétamine CD36 Transporteur d'acides gras
CD68 Marqueur de macrophage
CETP Protéine de transfert des esters de cholestérol
CRP Protéine C réactive
CT Tomodensitomètre à rayons X
db Gène du récepteur de la leptine DGAT Diacylglycérol acyltransférase DIO Diète riche en graisse et en sucre
ERK1/2 Kinase régulée par un signal extracellulaire FABP Protéine de liaison aux acides gras
FIAF Facteur adipeux induit par le jeûn FSV Fraction stroma-vasculaire
GAPDH Glycéraldehyde-3-phosphate déshydrogénase GLUT Transporteur de glucose
GPCR Récepteur couplé à la protéine-G HADH Hydroxyacyl-CoA déshydrogénase
HDL Lipoprotéine de haute densité HGF Facteur de croissance hépatique
HK Héxokinase
HMW 16-mères d’adiponectine de 400 à 600 kDa
HSL Lipase hormono-sensible
HyperapoB Hyperapolipoprotéine B ICV Intra-cérébro-ventriculaire
IGF-1 Facteur de croissance ressemblant à l’insuline-1
IL Interleukine
IL-1 Interleukine-1
IL-6 Interleukine-6
IMC Indice de masse corporelle IMCL Lipides intramyocellulaires IR Récepteur de l'insuline
IRS Substrat du récepteur de l'insuline
JAK Janus kinase
KC Chimiokine dérivée des kératinocytes KO Annulation complète d’un gène LCR Liquide céphalorachidien LDL Lipoprotéine de faible densité LHA Aire hypothalamique latérale LMW Homotrimères d’adiponectine
LPL Lipoprotéine lipase
LPS Lipopolysaccharide
mTOR La cible de la rapamycine chez les mammifères MAPK Protéine kinase activée par les signaux mitogènes MCH Hormone concentrant la mélanine
MCP-1 Protéine chimiotactique des monocytes 1 MIF Facteur inhibiteur de migration macrophagique MMW Hexamères d’adiponectine de 180 kDa
MPOC Maladie pulmonaire obstructive chronique Nampt Nicotinamide phosphoribosyltransférase NAD Nicotinamide adénine dinucléotide NEAT Activités physiques non programmées NGF Facteur de croissance de nerf (neurone) NFκB Facteur nucléaire
NO Monoxyde d'azote
NPY Neuropeptide Y
ob Gène de la leptine
ob/ob Souris déficiente en leptine
ob/obC3KO Souris déficiente en leptine et en ASP OMS Organisation mondiale de la santé
PAI-1 Inhibiteur d’activation de plasminogène-1 PBEF Facteur stimulant les colonies de cellules Pre-B PDE3B Phosphodiestérase 3
PEPCK Phosphoénolpyruvate carboxykinase
PGE2 Prostaglandine E2, PGI2 Prostaglandine I2,
PGF2-α 8-iso-prostaglandine F2-α PHOS Glycogène phosphorylase PI3K Phophatidylinositol-3 kinase
PKA Protéine kinase A
PKB/AKT Protéine kinase B/AKT
PKC Protéine kinase C
PLIN Périlipine A
POMC Neuropeptide pro-opiomélanocortine
PPAR Récepteur activé par les proliférateurs des peroxysomes PVN Noyau paraventriculaire
RIIα Sous-unité régulatrice de la PKA RAS Système rénine-angiotensine RBP-4 Protéine de liaison au rétinol-4 ROS Dérivés réactifs de l'oxygène
SAOS Syndrome d’apnées obstructives du sommeil SIRS Syndrome de réponse inflammatoire systémique
SNC Système nerveux central
SOCS3 Suppresseurs du signalement des cytokines
SOH Syndrome obésité-hypoventilation, syndrome hypoventilation alvéolaire
STAT3 Facteurs de transcription
T3 Triiodothyronine
TA Tissu adipeux
TAB Tissu adipeux blanc
TASC Tissu adipeux sous-cutané TAV Tissu adipeux viscéral
TEP Tomographie d'émission par positrons
TF Facteur de tissu
TG Triglycéride
TGF-β Facteur de croissance transformant-beta TNF-a Facteur de nécrose tumorale-a
TSH Hormone relâchant la thyrotropine
UCP Protéine découplante
VEGF Facteur de croissance de l'endothélium vasculaire VLDL Lipoprotéine de très basse densité
À ma famille, Marie-Josée et mes amis
pour m’avoir toujours appuyé dans mes projets
Remerciements
« Derrière chaque étudiant chercheur, se trouve une formidable équipe de recherche ».
Il m’est difficile de croire que tous les travaux réalisés au cours d’un projet de doctorat sont le fruit du labeur d’une seule personne. C’est pourquoi, j’aimerais profiter de ce chapitre pour remercier les personnes qui ont été impliquées de près ou de loin dans mes travaux de recherche.
Mes premiers remerciements vont à ma superviseure la Dre Katherine Cianflone grâce à qui je me suis initié à la recherche scientifique. Merci Katherine pour tant de patience à mon endroit, pour le partage généreux de tes connaissances, pour les discussions et les sorties lors des conférences, pour ta porte ouverte et, surtout, pour ta confiance. Merci beaucoup, Katherine!
Tout au long de mes travaux de recherche, j’ai eu la chance et surtout le plaisir de côtoyer des chercheurs de renommée internationale. Je tiens grandement à remercier les Drs Richard et Deshaies pour leurs nombreuses suggestions visant à améliorer mes travaux de recherche et les meilleurs endroits à visiter lorsqu’en conférence à l’extérieur du pays. Messieurs, le centre de recherche ne serait définitivement pas le même sans vous deux. Je ne peux écrire ce paragraphe sans remercier le Dr Patrick Schrauwen et son équipe pour leur accueil convivial lors de mon stage de perfectionnement à Maastricht. Des remerciements vont également à la Dre Timofeeva, au Dr Denis Joanisse, au Dr Buteau ainsi qu’au Dr Laplante, pour leur aide précieuse dans l’élaboration des diverses expériences composant mon projet de doctorat. Vos commentaires judicieux et votre expérience m’ont permis de m’engager avec confiance dans un axe de recherche peu connu de notre laboratoire. Merci à vous tous!
Je souhaite également souligner ma reconnaissance envers tous mes collègues. Tout d’abord, un gros merci à Sabina pour ta présence énergique, que ce soit lors de mes débuts dans un nouveau labo à Québec ou à Maastricht. Sab, tu as été une collègue fantastique sans qui bien de ces travaux de recherche n’auraient pas eu lieu et une amie merveilleuse. Je te serai toujours reconnaissant pour ton aide et ton soutien moral lors de mon séjour aux Pays-Bas. Marc, que dire de plus que wow! (peut-être pourde?), merci beaucoup pour ton
aide précieuse au labo, ton appui dans les moments les plus difficiles. Je me souviendrai des nombreux soupers au mexicain et des multiples pré-beer au BDL. Sans ta touche magique, le labo n’aurait jamais eu d’ASP et moi, pas de projet. Merci pour tout. Une mention spéciale pour Danny et Alexandre, sans qui la portion « labo » n'aurait pas été aussi agréable, efficace et stimulante. Les gars, merci de votre amitié hors labo, nos parties de cartes et nos soirées bien arrosées ont aussi fait partie de l’expérience. Mille mercis aux autres membres de l’équipe de recherche du Dre Cianflone, soit Huiling, Abhishek, Pierre, Mercedes, Reza et Pegah. Les agréables discussions, rencontres ainsi que les nombreuses activités de groupe ont contribué à la formation d'un environnement social où il fut plaisant de travailler. Merci aux professionnels de recherche, Julie, Yves et Pierre, puisque votre présence, plus que rassurante en laboratoire, fait en sorte que bien des expériences se terminent sur une bonne note plutôt que par un désastre. Enfin, un gigantesque merci à mes amis de l’animalerie : Justin, Sébastien, Nicolas, Caroline, Geneviève, Jennifer et Audrey. Amis et amies, aucun de ces articles n’aurait été publié sans votre expertise et votre professionnalisme, merci de tout cœur.
À mes amis de Kin, qui au cours des 13 dernières années ont su s’accommoder de mon mode de vie étudiante, merci à vous tous et toutes pour votre appui moral, votre amitié sans bornes et votre compréhension envers ma phobie pour le vrai monde du travail, je vous adore!!
To Lauren, Heather, Ralph, Florence, Boris, Bart and the rest of the Maastricht crew, heel erg bedankt!! Meeting all of you made my time in Maastricht a wonderful experience and I will cherish those memories forever. May our life paths cross again.
Je remercie les Instituts de recherche en santé du Canada (IRSC) et le Fonds de recherche en santé du Québec (FRSQ) pour le soutien financier accordé, soutien qui a servi à m’épauler au cours de mes études de troisième cycle.
Cette section ne pourrait se terminer sans l’expression de ma reconnaissance envers les membres de ma famille. En premier lieu, à mes parents, Hélène et Jean-Yves pour leur support quotidien et leur amour infini sans lesquels je ne serais pas qui je suis aujourd’hui. Merci de m’avoir poussé à apprendre. Merci surtout de toujours m’avoir encouragé à faire mon propre chemin. Merci à mes frères, Jean-François et Michel, vos encouragements tout
au long de mon cheminement ont été très appréciés et sachez que je suis très fier de vous deux.
Finalement, merci Marie-Josée : tu es entrée dans ma vie dans un moment professionnel difficile et tu as su me donner tout l’amour et le soutien moral dont j’avais besoin pour compléter cette aventure exigeante mais combien exaltante qu’est l’écriture d’une thèse, merci darling.
Avant-propos
Le présent ouvrage est déposé à la Faculté des études supérieures de l'Université Laval pour l'obtention du diplôme de Philosophiae Doctor ès Sciences (Ph.D.). Globalement, cette thèse porte sur le rôle de la voie ASP/C5L2 sur le métabolisme énergétique. Les analyses rapportées dans cet ouvrage s'attardent spécifiquement à trois aspects de la voie ASP/C5L2 : son rôle dans le contrôle de la prise alimentaire, son rôle dans l’utilisation des acides gras comme substrat énergétique par le muscle squelettique et l’implication de la voie ASP/C5L2 dans le phénotype des souris déficientes en C5aR. La thèse comprend une introduction générale rédigée en français portant sur les mécanismes impliqués dans le développement de l’obésité, le rôle du tissu adipeux blanc et brun dans le métabolisme énergétique, ainsi qu’une description du rôle de l’ASP et de son récepteur, le C5L2. Les chapitres 1 à 3 forment le corps de l'œuvre et exposent avec minutie les travaux de recherche accomplis pour cette thèse. Ces chapitres sont sous forme d'articles scientifiques rédigés en anglais, tels qu'ils ont été présentés aux éditeurs lors de leur publication.
Chapitre 1
Ce travail, réalisé sous la direction de la Dre Katherine Cianflone, a été rendu possible grâce à la collaboration de Danny Gauvreau (étudiant au doctorat avec la Dre Katherine Cianflone), de Fun-Qun Tom (étudiante à la maîtrise avec la Dre Katherine Cianflone) de la Dre Elena Timofeeva (CRIUCPQ) et du Dr Denis Richard (CRIUCPQ). La récolte des données effectuée par Danny Gauvreau et Fun-Qun Tom (analyse plasmatique) de même que la collaboration technique de Marie-Claude Roy et d’Anne-Marie Poulin (pour l’hybridation in situ) représentent des facteurs déterminants qui ont conduit à la publication du présent article. L'utilisation de la chambre métabolique ainsi que les chirurgies d’insertion de canules a été rendue possible grâce au Dr Denis Richard et son groupe de techniciens en santé animale (Nicolas Thiboutot-Gagnon, Caroline Gagnon et Geneviève Guèvremont). De plus, les docteurs Richard et Timofeeva ont grandement alimenté les discussions lors de l'écriture du manuscrit. Cette étude implique mon entière participation
quant à la réalisation de l'ensemble des étapes menant à la publication de cet article, soit la planification, l'exécution du protocole, la mesure des variables plasmatiques et tissulaires et l'analyse statistique. À la suite des corrections de la directrice, l'article a été révisé par l'ensemble des coauteurs et a été soumis aux éditeurs de la revue American Journal of
Physiology : Endocrinology and Metabolism. Cet article a été publié en juillet 2011.
Chapitre 2
Ce travail, réalisé sous la direction des Drs Katherine Cianflone et Patrick Schrauwen, a été rendu possible grâce à la collaboration de la coauteure Dre Sabina Paglialunga (stagiaire postdoctorale avec le Dr Patrick Schrauwen), de Ruth C. Meex et de Esther Phielix (étudiantes au doctorat avec le Dr Patrick Schrauwen), de Gert Schaart et de Esther Moonen-Kornips, (assistants de recherche du Dr Schrauwen), du Dr Joris Hoeks (professionnel de recherche du Dr Schrauwen) et du Dr Matthijs KC. Hesselink. La récolte de données par Ruth C. Meex et Esther Phielix (études humaines) de même que la collaboration technique de Gert Schaart et Esther Moonen-Kornips (pour la coloration Oil red O et l’immuno-bavardage) représentent des facteurs déterminants qui ont conduit à la publication du présent article. L'utilisation de l’oxygraph (OROBOROS, Oxygraph-2k) pour l’étude de la respiration mitochondriale à haute résolution a été rendue possible grâce au Dr Patrick Schrauwen. De plus, les docteurs Schrauwen, Hoeks et Hesselink ont généreusement contribué aux discussions lors de l'écriture du manuscrit. Cette étude implique mon entière participation et celle du Dre Paglialunga quant à la réalisation de l'ensemble des étapes menant à la publication de cet article, soit la planification, l'exécution du protocole, la mesure des variables plasmatiques et tissulaires et l'analyse statistique. À la suite des corrections des directeurs, l'article a été révisé par l'ensemble des coauteurs et a été soumis aux éditeurs de la revue PLOS ONE. Cet article a été publié en février 2013.
Chapitre 3
Ce travail, réalisé sous la direction de la Dre Katherine Cianflone, a été rendu possible grâce à la collaboration du Dr Abhishek Gupta (stagiaire postdoctoral avec la Dre Cianflone), d’Alexandre Fisette (étudiant au doctorat avec la Dre Cianflone), de Marc Lapointe (assistant de recherche du Dre Cianflone), de la Dre Huiling Lu (stagiaire
postdoctorale avec la Dre Cianflone), de Bao Lu et Norma Gerard (assistants de recherche du Dr Gerard) et des Drs Denis Richard et Craig Gerard. La mesure de l’ARN messager (PCR en temps réel) a été réalisée par le Dr Abhishek Gupta. Alexandre Fisette a contribué étroitement à la récolte des échantillons et à la mesure de certains paramètres, en plus de s'occuper des animaux. De plus, il a apporté une aide technique et conceptuelle précieuse lors de la réalisation des protocoles. La récolte de données par la Dre Huiling Lu (analyse plasmatique) de même que la collaboration technique de B. Lu et N. Gerard (pour les animaux modifiés génétiquement) représentent des facteurs déterminants qui ont conduit à la publication du présent article. L'utilisation de la chambre métabolique a été rendue possible grâce au Dr Denis Richard et la disponibilité d’animaux transgéniques est attribuable au Dr Gerard. De plus, les docteurs Richard et Gerard ont été très impliqués dans les discussions entourant l'écriture du manuscrit. Cette étude implique mon entière participation quant à la réalisation de l'ensemble des étapes menant à la publication de cet article, soit la planification, l'exécution du protocole, la mesure des variables plasmatiques et tissulaires et l'analyse statistique. À la suite des corrections des directeurs, l'article a été révisé par l'ensemble des coauteurs et a été soumis aux éditeurs de la revue PLOS ONE. Cet article a été publié en mai 2013.
La discussion qui termine cette thèse résume les principaux résultats obtenus et discute des voies futures à explorer afin d'améliorer la compréhension du rôle de la voie ASP/C5L2 dans le métabolisme énergétique.
Articles présentés
Les articles présentés dans cette thèse sont le résultat de mes travaux de doctorat effectués au cours des quatre dernières années sous la supervision de la Dre Katherine Cianflone. De la mise en place des protocoles à la rédaction finale des manuscrits, j’ai été impliqué à part entière dans chacune de ces études. De plus, lors de mes études supérieures, j'ai eu la chance de collaborer à plusieurs projets en plus de mes propres travaux de recherche. Ces collaborations avec mes collègues ou avec d'autres membres d’équipes de recherche ont conduit, ou conduiront prochainement, à la publication de manuscrits. L’énumération qui suit résume les articles provenant de tous mes travaux et collaborations. Les articles précédés d'un astérisque sont ceux qui seront présentés dans cette thèse.
* Roy C, Gupta A, Fisette A, Lapointe M, Poursharifi P, Richard D, Lu H, Lu B, Gerard N, Gerard C, Cianflone K. (2013) C5a Receptor Deficiency Alters Energy Utilization and Fat Storage. PloS ONE. 8(5): e62531. doi:10.1371/journal.pone.0062531, publié 7 mai 2013.
* Roy C*, Paglialunga S*, Schaart G, Moonen-Kornips E, Meex R, Phielix E, Hoeks J, Hesselink MKC, Cianflone K, Schrauwen P. C5L2-deficiency exacerbates diet-induced skeletal muscle fatty acid accumulation and insulin resistance. PLoS ONE. 2013;8(2):e57494. Epub 27 Février. 2013
* Roy C, Roy MC, Gauvreau D, Poulin AM, Tom FQ, Timofeeva E, Richard D, Cianflone K. Acute injection of ASP in the third ventricle inhibits food intake and locomotor activity in rats. Am J Physiol Endocrinol Metab. 2011 Jul;301(1): E232-41.
Poursharifi P, Lapointe M, Fisette A, Lu H, Roy C, Munkonda MN, Fairlie DP, Cianflone K.C5aR and C5L2 act in concert to balance immunometabolism in adipose tissue.Mol Cell Endocrinol. 2014 Jan 25;382(1):325-33.
Munkonda MN, Lapointe M, Miegueu P, Roy C, Gauvreau D, Richard D, Cianflone K. Recombinant acylation stimulating protein administration to C3-/- mice increases insulin resistance via adipocyte inflammatory mechanisms. PLoS ONE. 2012;7(10):e46883. Epub 2012 Oct. 8.
Gauvreau D, Roy C, Tom FQ, Lu H, Miegueu P, Richard D, Song WC, Stover C, Cianflone K. A new effector of lipid metabolism: complement factor properdin. Mol
Immunol. 2012 May;51(1):73-81. Epub 2012 Mar. 3.
Cianflone K, Paglialunga S, Roy C. Intestinally derived lipids: metabolic regulation and consequences-an overview. Atheroscler Suppl. 2008 Sep;9(2):63-8. Review.
Introduction
À l’échelle mondiale, le nombre de cas d’obésité a doublé depuis 1980 (OMS, 2008). L’obésité est devenue la pathologie nutritionnelle la plus fréquente dans les pays industrialisés. Sa progression est épidémique, elle apparaît de plus en plus précocement chez l’enfant et elle affecte une proportion d’individus de plus en plus importante, avec une augmentation plus marquée des obésités massives. Ses complications (diabète, maladies cardiovasculaires, cancer) ont un impact de plus en plus préoccupant sur la santé publique (OMS 2003; OMS 2007). En 2008, le surpoids touchait 1,5 milliard de personnes de 20 ans et plus, parmi lesquelles plus de 200 millions d’hommes et près de 300 millions de femmes étaient obèses (OMS, 2008). En 2010, le surpoids concernait près de 43 millions d’enfants de moins de cinq ans. Malgré tout, un facteur consolant existe : l’obésité est évitable.
1-Obésité
1.1-Définir le surpoids et l’obésité
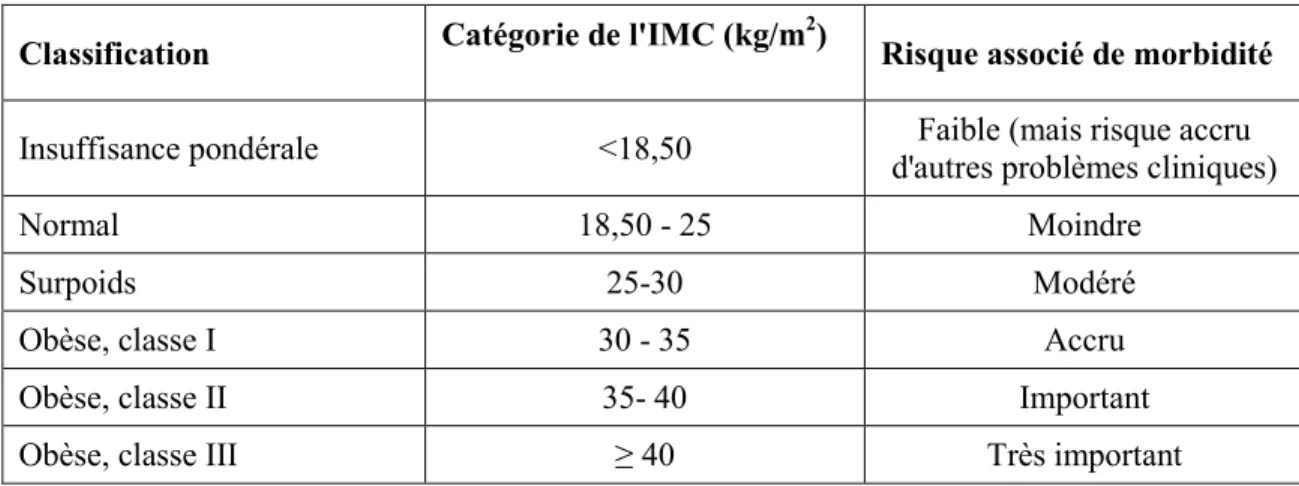
Le surpoids et l’obésité se définissent comme une accumulation anormale ou excessive de graisse corporelle entraînant des inconvénients pour la santé somatique, psychologique et sociale. Dans les années 90, l’OMS décida de se baser sur l’indice de masse corporelle (IMC), créée par Adolphe Quetelet au milieu du XIXe siècle et qui se calcule de la manière
suivante : poids (kg)/taille (m2). Depuis, cet indice sert de mesure universelle dans l’évaluation du surpoids et de l’obésité. Il permet de définir la charge pondérale des patients comme insuffisante (IMC< 18,5), normale (18,5 <IMC< 25), surcharge (IMC> 25), ou excédentaire (IMC> 30) et d’y associer un risque d’apparition de morbidités (Tableau 1). L’obésité est donc définie par un IMC> 30 kg/m2.
Tableau 1. Classification du risque pour la santé en fonction de l'indice de masse corporelle (IMC)
Classification Catégorie de l'IMC (kg/m2) Risque associé de morbidité
Insuffisance pondérale <18,50 d'autres problèmes cliniques) Faible (mais risque accru
Normal 18,50 - 25 Moindre
Surpoids 25-30 Modéré
Obèse, classe I 30 - 35 Accru
Obèse, classe II 35- 40 Important
Obèse, classe III ≥ 40 Très important
Source : Adapté de Santé Canada. Lignes directrices canadiennes pour la classification du poids chez les
adultes. Ministre des Travaux publics et Services gouvernementaux du Canada, 2003.
L’IMC est sans contredit une mesure utile du surpoids et de l’obésité dans une population puisque, chez l’adulte, l’échelle est la même quels que soient le sexe ou l’âge du sujet. Toutefois, l’IMC donne une indication approximative, car il ne correspond pas forcément au même degré d’adiposité d’un individu à l’autre. C’est pourquoi, en pratique clinique courante, l’estimation de la masse grasse est basée sur des mesures anthropométriques telles que le poids, la taille, les plis cutanés et la circonférence de la taille (ANAES, 2003). Il est maintenant admis qu’indépendamment de l’IMC, l'excès de tissu adipeux au niveau de la cavité abdominale (obésité de type androïde, Figure 1) est beaucoup plus néfaste pour la santé qu’au niveau des cuisses et des hanches (obésité de type gynoïde, Figure 1).
Figure 1. La distribution régionale du tissu adipeux.L’obésité androïde, communément appelée obésité abdominale ou viscérale, se caractérise par une accumulation de tissu adipeux dans la région abdominale et comprend les dépôts sous-cutanés et les dépôts viscéraux. L’obésité gynoïde se caractérise par une accumulation préférentielle de tissu adipeux dans la région hanches-cuisses, qui constitue un dépôt sous-cutané.Modifié de conf-de-nutrition.blogs.fr
L'obésité androïde serait la forme la plus néfaste pour la prévalence du syndrome métabolique (Tableau 2)(Despres, J.P., 2006b). Dans cette optique, Yusuf et coll. (Dagenais, G.R., 2003) ont démontré qu'à l'intérieur de chaque catégorie d'IMC, une augmentation du rapport taille/hanches (indice pour estimer l’accumulation du tissu adipeux au niveau abdominal) est associée à une augmentation subséquente du risque d'infarctus du myocarde. De plus, il a été observé que les sujets obèses ayant une faible quantité de tissu adipeux viscéral ont une tolérance au glucose similaire à ceux de poids normal (Pouliot, M.C., 1992).
Récemment, l’étude transversale International Day for the Evaluation of Abdominal Obesity (IDEA) a démontré l'importance de la circonférence de la taille en tant que variable
associée au risque de maladies cardiovasculaires et de diabète de type 2 (Balkau, B., 2007). Ces résultats montrent qu’en plus d’une augmentation de la masse grasse, la distribution du tissu adipeux est un élément important dans la physiopathologie cardiovasculaire liée à l’obésité.
Tableau 2. Risques relatifs de complication métabolique en fonction du tour de taille
Genre Périmètre abdominal (cm) Risque de complication métabolique
Masculin >94 Accru
>102 Important
Féminin >80 Accru
>88 Important
En résumé, la classification des personnes en surpoids et obèses se fait actuellement par l’utilisation de ces deux mesures, soit IMC et rapport taille/hanches. Cependant, le calcul de l’IMC sert de référence préférée pour l’estimation de la prévalence de l’obésité au sein des populations.
1.2-Les causes de l’obésité et du surpoids
La cause fondamentale de l’obésité et du surpoids est un déséquilibre énergétique entre les calories consommées et dépensées (Figure 2). Au niveau mondial (OMS, 2008), on a relevé:
• une plus grande consommation d’aliments très caloriques, riches en graisses et en sucres, mais pauvres en vitamines, en minéraux et autres micronutriments, et; • une baisse de la pratique d’exercice physique en raison de la nature de plus en plus
sédentaire de nombreuses formes de travail, de l’évolution des modes de transport et de l’urbanisation.
Figure 2.La balance énergétique. Ce diagramme montre les principes fondamentaux de l’équilibre et de la régulation
énergétique. Il y a bilan énergétique positif lorsque l’apport calorique est supérieur à la dépense ce qui favorise la prise de poids. À l’inverse, un bilan énergétique négatif favorise la diminution des réserves de graisse de l’organisme et la perte de poids. Modifié de www.diet-ethique.toile-libre.org
De nombreuses études ont mis en évidence une régulation homéostatique du poids corporel qui permet de maintenir un poids relativement stable malgré des apports et des dépenses énergétiques variables (Berthoud, H.R., 2006). On pense que l’organisme se défend plus vigoureusement contre la dénutrition et la perte de poids qu’il ne le fait contre la surconsommation et la prise de poids. D'un point de vue quantitatif, le gain de poids est un processus se déroulant souvent sur plusieurs années. En effet, l’étude de Framingham a montré que la prise de poids moyenne sur une période de 18 ans est d’environ 10 kg (Lissner, L., 1991). De plus, le déséquilibre énergétique n'a pas besoin d'être important pour conduire à une obésité. En ce sens, des apports quotidiens dépassant de 5 % les dépenses annuelle pouvant aller jusqu’à 5 kg de masse grasse, suggérant qu'un faible déséquilibre entre l'apport et la dépense énergétique peut mener à une obésité (Jequier, E., 2002). Également, des forces sociétales et environnementales importantes influent sur l’apport et la dépense énergétiques et peuvent submerger les processus physiologiques mentionnés précédemment (Figure 3). La sensibilité des individus à ces forces est fonction de facteurs génétiques et biologiques tels que le sexe, l’âge et l’activité hormonale, sur lesquels ils ont peu ou pas de contrôle. On estime que les facteurs diététiques et l’inactivité physique sont les facteurs intermédiaires modifiables à travers lesquels les forces favorisant la prise de poids s’exercent.
Figure 3. Facteurs d’influence sur le poids. Bien que multifactorielles, les causes les plus souvent évoquées pour
expliquer l’émergence de l’obésité sont une prédisposition génétique associée à un mode de vie « inadapté », où les erreurs alimentaires occupent une place majeure, et où l’inactivité physique s’inscrit dans les habitudes journalières. Tiré de www.obesite.ulaval.ca
1.2.1 Dépense énergétique
La dépense énergétique est divisée en trois composantes : le métabolisme de repos, la thermogenèse et l’activité physique. Elle peut être influencée par des facteurs intrinsèques tels que la masse de tissu maigre, l’âge et le sexe ou par des facteurs extrinsèques tels que la prise alimentaire ou l’exposition au froid.
1.2.1.1 Le métabolisme de repos
Le métabolisme de repos est défini comme la dépense énergétique d’un sujet en état postprandial au repos à la neutralité thermique. La dépense énergétique représente la consommation d’énergie nécessaire pour assurer les fonctions vitales de l’organisme : travail des pompes membranaires nécessaires pour maintenir les gradients ioniques, renouvellement des constituants de l’organisme, travail cardiaque et respiratoire (Martin, C., 2006). Chez un individu ayant une activité physique moyenne, on estime que le métabolisme de repos représente environ 60 % de la dépense énergétique totale (Leibel, R.L., 1995). La contribution des divers organes à la dépense énergétique est très diversifiée,
la majeure partie du métabolisme de repos étant due à la dépense énergétique d’organes tels que le foie, le cerveau, le coeur, les reins et de celle des muscles au repos. Le métabolisme de repos varie principalement en fonction de la masse maigre et plus particulièrement de la masse musculaire, ce qui explique la grande variabilité interindividuelle (Johnstone, A.M., 2005). On estime généralement que la masse maigre peut expliquer 70 à 80 % de la variabilité du métabolisme de repos contre 2 % pour la masse grasse. Cependant, chez certains sujets fortement obèses, cette valeur peut atteindre 10 % pour le tissu adipeux (Bergouignan, A., 2006). Ainsi, les différences de métabolisme liées au sexe et à l’âge seraient en grande partie dues à des différences de masse maigre, moins importante chez la femme que chez l’homme, différences se résorbant avec l’âge. Des études chez l’obèse ont démontré que 75 % du poids gagné est dû au développement du tissu adipeux et 25 % à une augmentation de la masse maigre. On observe donc chez l’obèse une augmentation du métabolisme de repos, mais si l’on rapporte cette valeur à la masse maigre, aucune différence significative n’a pu être notée (Bandini, L.G., 1990; Treuth, M.S., 1998). En revanche, on constate chez l’obèse amaigri, une diminution du métabolisme de repos qui pourrait être à l’origine de la reprise de poids fréquemment rencontrée après le régime (Lazzer, S., 2004).
1.2.1.2 La thermogenèse adaptative
La thermogenèse adaptative correspond à l’énergie dissipée sous forme de chaleur en réponse à des changements environnementaux tels que le froid, le régime, le stress ou l’administration de substances stimulant le métabolisme (ex. : caféine, nicotine)(Lowell, B.B., 2000). Parmi ces facteurs, la prise alimentaire est la plus importante. L’effet thermique des repas représente environ 10 % des dépenses énergétiques quotidiennes (Leibel, R.L., 1995). Cette thermogenèse postprandiale comprend la thermogenèse « obligatoire » et « facultative ». La thermogenèse obligatoire correspond à la transformation et à la mise en réserve des nutriments. Le métabolisme du glucose en glycogène requiert 5 % de l’énergie contenue dans le glucose alors que la lipogenèse à partir de glucose en nécessite cinq fois plus. Le stockage des lipides dans le tissu adipeux sous forme de triglycérides demande environ 2 % de l’énergie ingérée sous forme de lipides. Quant à la transformation des protéines, elle nécessite 25 % de l’énergie des
protéines ingérées afin d’assurer la néoglucogenèse, l’uréogenèse et la stimulation de la synthèse protéique (Joosen, A.M., 2006). La thermogenèse postprandiale « facultative », quant à elle, intervient en condition de suralimentation. Elle permettrait à l’organisme de dissiper sous forme de chaleur l’énergie reçue en excès (Levine, J.A., 1999).
1.2.1.3 L’activité physique
L’activité physique est la composante la plus variable de la dépense énergétique totale, car elle dépend du mode de vie des individus. De manière générale, elle représente 20 à 30 % de la dépense énergétique totale, mais chez les individus très sportifs ou effectuant un travail de force, elle peut atteindre 60 % de la dépense énergétique totale (Rising, R., 1994). Au cours des cent dernières années, l’utilisation de machines pour les travaux agricoles, domestiques et industriels a entraîné une diminution de l’activité physique. On estime que la dépense énergétique journalière a diminué de 600 kcal entre les années 1920 et 1990. Les travaux de Prentice défendent l’idée que la diminution de l’activité physique pourrait être la cause de l’obésité puisque l’augmentation de la prévalence de l’obésité s’accorde avec un mode de vie de plus en plus sédentaire (Prentice, A.M., 1995), sans oublier qu’une activité physique volontaire joue un rôle primordial dans la prévention de l’obésité (Warburton, D.E., 2006). En 1999, une nouvelle forme de dépense énergétique jusque-là ignorée a été identifiée. Il s’agit de la thermogenèse induite par les activités physiques non programmées (Non exercice activity thermogenesis : NEAT). Ces activités physiques réunissent l’ensemble des mouvements non volontaires tels que le maintien de la posture, la position debout ou encore les mouvements liés à la nervosité. Levine et son équipe ont récemment montré que la dépense énergétique liée à la NEAT pouvait expliquer la différence de poids entre des individus sédentaires minces et des obèses (Levine, J.A., 2005). En effet, il semblerait que les sujets obèses passent deux heures de plus en position assise que les sujets minces, ce qui représente une différence de dépense énergétique de plus de 350 kcal/jour. Un bilan énergétique positif aboutit à une inévitable augmentation du poids corporel principalement au niveau du tissu adipeux. Le tissu adipeux représente la réserve énergétique la plus importante parce qu’il permet d’emmagasiner l’excédent énergétique sous le poids le plus faible et sous la forme la plus dense. En effet, on estime qu’un gramme
de graisse contient six fois plus d’énergie qu’un gramme de glycogène. Chez un sujet mâle de 70 kg, les triglycérides représentent environ 12 kg de son poids total. L’équivalent énergétique en glycogène impliquerait un poids corporel plus élevé de 55 kg.
1.2.2 Apport énergétique
Au-delà de l’activité ou l’inactivité physique, la cause comportementale la plus étudiée susceptible d’influencer le poids corporel et ainsi le risque de faire de l’embonpoint et de l’obésité est l’alimentation. Bien que la majeure partie des données accessibles porte essentiellement sur des rapports de corrélation, l’homogénéité des données dans leur ensemble fait ressortir l’importance de bonnes habitudes alimentaires et l’accès à des aliments sains comme éléments clés associés à la prévention de l’obésité au niveau de la population. Un bon nombre d’études a permis d’observer un rapport entre une faible consommation de fruits et légumes (un indicateur d’alimentation pauvre) et l’obésité (Bauman, A., 2005). Également, des études de modélisation de l’apport énergétique canadien et des niveaux de dépense menées entre 1976 et 2003 ont révélé un fort lien entre la prévalence de l’obésité grandissante et la consommation croissante de calories, cette dernière étant composée par sept groupes d’aliments (huiles à salade, farine de blé, boissons gazeuses, saindoux, riz, poulet et fromage) (Slater, J., 2009). Chez les enfants et les adolescents, les facteurs familiaux et environnementaux peuvent être associés à des choix et à des comportements alimentaires particuliers (Janssen, I., 2006; Minaker, L.M., 2006; Veugelers, P., 2008). Par exemple, le fait de prendre des collations ou de grignoter en regardant la télévision (Dubois, L., 2008; Liang, T., 2009), la consommation de boissons sucrées entre les repas (Dubois, L., 2007), et l’absence de petit-déjeuner (Dubois, L., 2009) ont été associés à un accroissement du risque d’obésité chez les enfants et les jeunes. De plus, une étude menée auprès d’enfants fréquentant l’école intermédiaire a permis de constater qu’une fréquence plus élevée de repas pris en famille était associée à une diminution de la consommation de boissons gazeuses, à une assiduité accrue au petit-déjeuner, à une diminution des problèmes d’obésité et à une plus grande efficacité de la saine alimentation à domicile et durant le temps passé avec les camarades (Woodruff, S.J., 2009).
Pour certains, l’objectif principal de la prise en charge nutritionnelle des sujets obèses reste la réduction pondérale. Les méthodes vont de la restriction calorique plus ou moins importante à des modifications de la composition de l’apport alimentaire. Ainsi, la restriction calorique représente le moyen le plus utilisé (surtout au début de la prise en charge) dans la lutte contre l’excès de poids et de la masse grasse chez les sujets obèses (Strychar, I., 2006). De manière générale, la ration calorique dans ce type de régime est calculée en fonction des apports habituels et de l’estimation des dépenses énergétiques, avec une réduction de l’ordre de 25 à 30 % par rapport aux apports antérieurs. Si les régimes restrictifs permettent effectivement d’obtenir une perte de poids sur un temps relativement court, l’approche diététique doit surtout se focaliser sur la modification du comportement alimentaire pour atteindre une alimentation conforme aux recommandations nutritionnelles (PNNS, 2001).
En contre-partie, il y a beaucoup d’études sur les comportements alimentaires qui montrent une reprise de poids après une diète de réduction pondérale, laissant croire que de manière générale la restriction alimentaire cognitive stricte peut augmenter le risque de reprise de poids perdu (Wu, T., 2009). Enfin, certains cherhceurs comme le Dr Després, suite à son étude Synergie (Borel, A.L., 2012a; Borel, A.L., 2012b), préfère, grâce à une intervention nutritionnelle, cibler la réduction du tour de taille et de la graisse viscérale, pour améliorer la qualité de vie des patients obèses.
1.3-Les pathologies reliées au surpoids et à l’obésité
Un IMC élevé, et, encore plus, un tour de taille imposant sont tous deux d’importants facteurs de risque de maladies chroniques. Une des premières hypothèses concernant cette relation fut émise, il y a plusieurs siècles, par Hippocrate (v.460-377 av. J.-C.) dans l’un de ses aphorismes : « l’anomalie de la corpulence n’est pas seulement une maladie en
elle-même, elle prédispose aussi à la survenue d’autres maladies ». Ainsi, quatre siècles avant
Jésus-Christ, l’obésité était déjà reconnue comme un désordre médical pouvant s’accompagner de nombreuses comorbidités. Selon l’OMS, le surpoids et l’obésité sont le cinquième facteur de risque de décès au niveau mondial. Au moins 2,8 millions d’adultes en meurent chaque année. En outre, le surpoids et l’obésité seraient responsables de plus de
30 % de la charge du diabète, de 23 % de la charge des cardiopathies ischémiques et de 7 à 41 % de la charge de certains cancers (OMS, 2010).
1.3.1 Diabète de type 2
Le diabète de type 2 s’installe progressivement dans l’histoire de l’individu obèse (Fery, F., 2005; Hu, F.B., 2001; Scheen, A.J., 2000; Scheen, A.J., 2005) (Figure 4). La présence d’antécédents familiaux contribue à l’apparition, plus ou moins précoce, de l’intolérance au glucose. L’obésité, spécialement celle caractérisée par une adiposité intra-abdominale, est associée à une augmentation des concentrations des acides gras libres circulants qui exercent un effet négatif sur la sensibilité à l’insuline au niveau de plusieurs tissus, dont le muscle et le foie. Les acides gras libres augmentent la synthèse hépatique du glucose en stimulant la néoglucogenèse, ils diminuent l’extraction de l’insuline et ils augmentent la production des lipoprotéines de très basse densité (VLDL) par le foie. La résistance à l’insuline passe inaperçue au début et ne sera diagnostiquée que par la présence d’une hyperinsulinémie, d’abord en postprandial, puis à l’état basal. Le diabète de type 2 apparaît lorsque la sécrétion pancréatique d’insuline devient insuffisante par rapport à la résistance des tissus cibles, pour atteindre une normoglycémie (Fery, F., 2005; Hu, F.B., 2001; Scheen, A.J., 2000; Scheen, A.J., 2005).
Figure 4. Illustration des mécanismes liant l’obésité au diabète de type 2. Le diabète de type 2 est une maladie
évolutive qui survient à la suite de cercles vicieux impliquant la glucotoxicité et la lipotoxicité en passant par la diminution de la tolérance au glucose (IGT). Modifié de M. RORIVE ET COLL. Rev. Med. Liège 2005.
Les données du Third National Health and Nutrition Heart Study Survey (NHANES III) montrent une association directe entre l’IMC et la prévalence du diabète de type 2 chez l’adulte (Harris, M.I., 1998). Il est clairement établi que le risque de développer un diabète de type 2 (diabète non insulinodépendant) augmente avec l’IMC et diminue avec la perte de poids, seule ou combinée à l’activité physique (Lundgren, H., 1989; Schmidt, M.I., 1992; Williams, K.V., 2000).
On estime à l’heure actuelle que 75 % des patients diabétiques de type 2 sont obèses et que, pour une valeur d’IMC supérieure ou égale à 30, comparé à un IMC inférieur à 30, le risque de diabète de type 2 est multiplié par dix chez la femme et par huit chez l’homme. L’adiposité viscérale en particulier augmente le risque d’hyperinsulinémie et d’intolérance au glucose (Kaye, S.A., 1991). Finalement, le diabète de type 2, auparavant considéré comme le diabète de l’adulte d’âge mûr, est maintenant retrouvé chez l’enfant et l’adolescent et y est aussi fréquent que celui de type 1 (Alberti, G., 2004; Rosenbloom, A.L., 1999).
1.3.2 Autres perturbations endocriniennes et métaboliques
Il se trouve que l’on observe un profil hormonal modifié chez les obèses, surtout parmi ceux arborant une accumulation de graisse intra-abdominale (Seidell, J.C., 1990). La sensibilité à l’insuline varie amplement entre les personnes mais, de manière générale, la résistance à l’insuline est associée à l’obésité particulièrement chez les personnes présentant beaucoup de tissu adipeux abdominal (Kissebah, A.H., 1989). De plus, cette résistance est systématiquement observée chez les personnes ayant un IMC> 40, ce qui a porté certains scientifiques à émettre l’hypothèse que la résistance à l’insuline serait un mécanisme d’adaptation à l’obésité afin de limiter le développement du tissu adipeux (Eckel, R.H., 1992; Kissebah, A.H., 1989). Enfin, chez les femmes, l’augmentation du poids corporel et l’hyperinsulinémie sont souvent liés à une irrégularité des cycles menstruels (Frontini, M.G., 2003), alors que l’obésité intra-abdominale est souvent associée au syndrome des ovaires polykystiques (Barber, T.M., 2006).
Une des perturbations métaboliques observée est la dyslipidémie où les taux de triglycérides plasmatiques et de lipoprotéines de faible densité (LDL) sont élevés et les taux de lipoprotéines de haute densité (HDL) sont bas. Ce profil métabolique est souvent observé chez les personnes obèses présentant une accumulation importante de tissu adipeux abdominal (Despres, J.P., 1990). Le risque de dyslipidémie augmente progressivement à partir d’un IMC de 21 (Haslam, D.W., 2005).
De nombreuses autres perturbations métaboliques sont rencontrées lors de l’obésité et ont été regroupées sous le nom de « syndrome métabolique » (Grundy, S.M., 2005). Comme passée en revue par Blackburn et collégues (Blackburn, P., 2009), plusieurs organsimes ont proposés des critères diagnostiques pour le syndrome métabolique, créant ainsi une variété d’outils disponible pour cibler les patients à risque.
En général, la définition clinique du syndrome métabolique repose sur six paramètres, où la présence de trois suffit à poser le diagnostic :
une obésité centrale : tour de taille > 102 cm chez les hommes et > 88 cm chez les femmes (varie selon le groupe ethnique),
un faible taux de cholestérol de type HDL : HDL < 40 mg/dl chez les hommes et <50 mg/dl chez les femmes,
une hypertriglycéridémie : triglycérides > 150 mg/dl,
une élévation de la pression sanguine : tension artérielle > 130-85 mm Hg, une diminution de la tolérance au glucose,
une résistance à l’insuline.
1.3.3 Maladies cardiovasculaires
Dès 1980 des études épidémiologiques importantes reconnaissent le lien entre l’obésité et les maladies cardiovasculaires (Eckel, R.H., 1997). De fait, l’obésité augmente les risques cardiovasculaires entres autres en favorisant les facteurs de risques tels que l’hypertension, les dyslipidémies, la résistance à l’insuline, mais également par ses effets sur de nouveaux types de facteurs de risque tels que les marqueurs de l’inflammation (ex. : protéine C réactive) ou de la thrombose (ex. : fibrinogène) (Berk, B.C., 1990; de Maat, M.P., 1996; Yudkin, J.S., 1999).
Les maladies cardiovasculaires sont responsables d’une large part de la mortalité dans les pays industrialisés et leur incidence augmente dans les pays en voie de développement. De nombreuses études ont démontré une relation linéaire entre surpoids et problèmes cardiaques. En plus de la quantité de masse grasse, il faut prendre en compte le profil de distribution de cette masse. En effet, le tissu adipeux localisé dans la partie supérieure du corps (la taille, l’abdomen) apparaît être un indicateur de qualité dans l’apparition de risque cardiovasculaire (Balkau, B., 2007; Despres, J.P., 2006a; Yusuf, S., 2005).
1.3.4 Problèmes respiratoires
Les principales complications respiratoires chez le sujet obèse sont le syndrome obésité-hypoventilation (SOH, aussi connu sous le nom de syndrome obésité-hypoventilation alvéolaire ou le syndrome Pickwickien) (BICKELMANN, A.G., 1956), le syndrome d’apnées
obstructives du sommeil (SAOS), l’hypertension artérielle pulmonaire et la maladie pulmonaire obstructive chronique (MPOC). Alors que 3 à 4 % des adultes souffrent de SAOS (Young, T., 1993), la prévalence de cette dernière pourrait dépasser 50 % dans le cas d’obésité massive (Resta, O., 2001). Pour sa part, le SOH est observé chez un peu moins de 10 % des obèses sévères (Weitzenblum, E., 2008). Pour ce qui est de la MPOC, la littérature suggère que 20 à 50 % des sujets souffrant de celle-ci sont également obèses (Eisner, M.D., 2007; Steuten, L.M., 2006).
1.3.5 Cancers
De plus en plus, les études scientifiques démontrent qu’un excès de poids augmente le risque de développer un cancer. Effectivement, chez l’homme, l’obésité est associée à un risque élevé de cancer de la prostate, colorectal et du rein (Chow, W.H., 2000). Les résultats vont dans le même sens chez la femme, où les risques de développer un cancer de l’endomètre, des voies biliaires, du col de l’utérus, des ovaires et du sein après la ménopause sont plus élevés chez les sujets obèses (Schindler, A.E., 1997). On estime que 34 à 56 % des cancers de l’endomètre et plus de la moitié des cancers du sein postménopause sont attribuables à une augmentation du poids corporel (IMC> 29) (Ballard-Barbash, R., 1996). L’augmentation de l’incidence de ces cancers chez les personnes obèses est plus importante en présence d’une forte accumulation de tissu adipeux intra-abdominal (Schapira, D.V., 1994).
1.3.6 Stéato-hépatite non alcoolique
La prévalence de la stéato-hépatite non alcoolique augmente rapidement dans les pays développés. Elle survient habituellement dans un contexte particulier associant obésité, diabète de type 2 et hypertriglycéridémie (Schwimmer, J.B., 2006; Seppala-Lindroos, A., 2002; Wanless, I.R., 1990). Sheth et ses collaborateurs ont montré que plus de 70 % des patients présentant une stéato-hépatite non alcoolique sont en surpoids (Sheth, S.G., 1997). Même si au départ cette maladie est généralement considérée comme bénigne, elle peut, dans certains cas, conduire à l’apparition d’une cirrhose ou d’une insuffisance hépatique pouvant même nécessiter une transplantation. L’augmentation des dépôts graisseux
viscéraux, des acides gras libres, et de l’hyperinsulinémie engendrée par l’obésité semble être factorielle de ce processus (Saito, T., 2007; Sheth, S.G., 1997). Finalement, la stéato-hépatite non alcoolique est une maladie qui semble être réversible par une perte de poids (Eriksson, S., 1986).
2-Tissus adipeux
Le tissu adipeux (TA) joue un rôle primordial dans l’homéostasie métabolique de l’organisme. Chez les mammifères, deux types de tissus adipeux sont classiquement distingués morphologiquement et fonctionnellement (Figure 5) : le tissu adipeux blanc (composé majoritairement d’adipocytes blancs) et le tissu adipeux brun (composé majoritairement d’adipocytes bruns) (Prunet-Marcassus, B., 2006).
Figure 5. Adipocyte blanc vs adipocyte brun. Outre leur noyau (N), l’adipocyte blanc et l’adipocyte brun présentent une
morphologie bien distincte l’une de l’autre. L’adipocyte blanc contient une seule vacuole lipidique (LV) et quelques mitochondries (M) alors que l’adipocyte brun contient plusieurs vacuoles lipidiques et un nombre très élevé de mitochondries. Modifié de www.sportsci.org
Le tissu adipeux blanc constitue le tissu adipeux principal chez l’homme adulte. Outre son rôle d’isolant thermique et mécanique, c’est la principale réserve énergétique de l’organisme (Prunet-Marcassus, B., 2006). Le rôle du tissu adipeux blanc est de réguler la balance énergétique en stockant l’énergie en surplus sous forme de triglycérides (TG) qui restent mobilisables pour les autres tissus en fonction des besoins métaboliques (Casteilla, L., 1994). Chez l’homme moyen de 70 kg, il va représenter 15 kg, soit 21 % de sa masse
corporelle. Ce pourcentage est augmenté chez la femme et au cours de l’obésité. Pour sa part, le tissu adipeux brun intervient essentiellement dans la thermogénèse (Casteilla, L., 1994).
2.1-Tissu adipeux brun
Tout récemment en 2011, grâce à l’utilisation couplée de la technique de tomographie d'émission par positrons (TEP) avec un tomodensitomètre à rayons X (CT), le tissu adipeux brun a été localisé chez l’humain adulte. D’un point de vue anatomique, on retrouve ce tissu dans deux types de dépôts : ceux des sites bien précis tels que les régions périrénale, cervicale ou encore interscapulaire et, ceux des sites diffus où le tissu adipeux brun coexiste avec des tissus de type adipeux blanc ou muscle squelettique (Ravussin, E., 2011).
Le tissu adipeux brun participe de manière active au phénomène de thermogenèse dite de non-frisson par opposition à la production de chaleur assurée par le frissonnement musculaire. La distribution efficace de la chaleur produite par ce tissu adipeux est assurée par une vascularisation importante. Les adipocytes bruns sont plus petits que les adipocytes blancs et possèdent plusieurs gouttelettes lipidiques ainsi que de nombreuses mitochondries (Figure 5). L’activité thermogénique du tissu est due à l’expression de la protéine découplante, UCP1 (Uncoupling protein 1) (Nicholls, D.G., 1999). Le rôle principal des mitochondries est d’assurer la phosphorylation oxydative. Ce processus se met en place grâce à des réactions qui ont lieu au niveau de la matrice mitochondriale et au niveau de la membrane interne. Le passage des protons de l’espace intermembranaire vers la matrice mitochondriale se fait via l’UCP1. L’énergie correspondante à cette force protomotrice est alors dissipée sous forme de chaleur (thermogenèse). Le contrôle de l’activité thermogénique des adipocytes bruns est sous la dépendance des récepteurs de type béta3-adrénergiques. En effet, au cours d’une exposition au froid, la noradrénaline libérée augmente le nombre de mitochondries, l’expression d’UCP1 et le nombre d’adipocytes bruns (Bukowiecki, L.J., 1986; Ricquier, D., 2002).
Bien qu'il soit possible maintenant de conclure que la dépense d'énergie reliée à l'exposition au froid peut contribuer à faire perdre du poids, la quantité d'énergie consommée est faible
si on la compare à toute autre forme d'activité physique (Ravussin, E., 2011). Les résultats de ces recherches sont toutefois encourageants. Il n'est pas exclu d'entrevoir un traitement de l’obésité ciblant l'activation métabolique des graisses brunes (Haas, B., 2012) accompagné d’exercices physiques et d’une bonne alimentation.
2.2-Tissu adipeux blanc
En 1965, Jean Vague et Robert Fenasse décrivaient deux types de tissus adipeux blancs selon leur localisation : le tissu adipeux viscéral (TAV) et le tissu adipeux sous-cutané (TASC) (Figure 6). Il a ainsi été observé que le tissu adipeux profond se répartit dans la cavité péritonéale et l’espace rétro-péritonéal. Dans la cavité péritonéale, on trouve le tissu adipeux viscéral composé de dépôts mésentériques, omentaux et perigonadaux. Ces dépôts représenteraient environ 20 % (chez l’homme) et 5 % (chez la femme) de la graisse corporelle totale (Lafontan, M., 2003). Dans l’espace rétro-péritonéal se trouvent les tissus adipeux périrénaux, pancréatiques et urogénitaux.
Il a été observé qu’une diminution du TAV améliore la sensibilité à l’insuline de l’organisme et induit le métabolisme du glucose. En effet, la masse de TAV est positivement corrélée avec l’intolérance au glucose, l’altération du niveau plasmatique de lipoprotéines, l’augmentation des concentrations de triglycérides et de cholestérol, l’hypertension et la dyslipidémie (Klein, S., 2004; Thorne, A., 2002). De plus, l’analyse des voies de signalisation de l’insuline dans les TA viscéral et sous-cutané humains montre que le TAV exprime des niveaux plus élevés de protéines spécifiques de la voie de signalisation de l’insuline et une plus grande sensibilité à l’insuline (Laviola, L., 2006). De fait, il a été démontré que le TAV est plus sensible à la perte de poids que le TASC, plus actif métaboliquement, plus lipolytique et produit plus d’adipokines (Park, H.S., 2005).
Le tissu adipeux sous-cutané quant à lui est divisé en deux couches soit : superficielle et profonde. Ce tissu est essentiellement localisé au niveau abdominal et fémoral. À l’inverse du tissu adipeux viscéral plus abondant chez l’homme, ce tissu semble plus répandu chez la femme avant la ménopause. Jean Vague a été le premier à distinguer une répartition des dépôts adipeux différente selon le sexe et à y attribuer des pathologies associées distinctes
(Vague, J., 1947; Vague, J., 1956). De cette répartition émergera le profil pomme/poire précédemment évoqué (voir 1.1 et Figure 1). Les dépôts sous-cutanés sont ceux qui subissent les plus importants changements de taille sans effet notable sur la sensibilité à l’insuline, le métabolisme du glucose ou le profil métabolique de l’organisme (Despres, J.P., 1989a; Despres, J.P., 1989b). Grâce à ces études, il apparaît très clairement que ces deux types de dépôts, par leurs activités sécrétoires et leurs réponses distinctes aux stimuli hormonaux, sont à l’origine des complications du surpoids et de l’obésité (Despres, J.P., 2006b; Lafontan, M., 2003).
Figure 6. Distribution anatomique du tissu adipeux blanc. Le tissu adipeux blanc, (TA blanc) est divisé en deux types
de tissus possédant des caractéristiques propres : le TA blanc sous cutané (TASC) et le TA blanc viscéral (TAV). Modifié de Anghel et Wahli, Cell Research (2007).
2.2.1 Emmagasinage de l’énergie
L’adipocyte blanc (Figure 5), siège principal de l’activité métabolique du TA est une cellule extensible possédant la particularité de pouvoir s’adapter en fonction des apports et des besoins énergétiques de l’organisme. Outre cette capacité de stockage des lipides, l’adipocyte blanc possède une activité sécrétrice de nombreux facteurs agissant de façon