Diversité microbienne associée au cycle du méthane
dans les mares de fonte du pergélisol subarctique
Thèse
Sophie Crevecoeur
Doctorat en biologie
Philosophiae doctor (Ph. D.)
Québec, Canada
© Sophie Crevecoeur, 2016
Diversité microbienne associée au cycle du méthane
dans les mares de fonte du pergélisol subarctique
Thèse
Sophie Crevecoeur
Sous la direction de :
Warwick F. Vincent, directeur de recherche
Connie Lovejoy, codirectrice de recherche
iii
Résumé
La fonte et l’effondrement du pergélisol riche en glace dans la région subarctique du Québec ont donné lieu à la formation de petits lacs (mares de thermokarst) qui émettent des gaz à effet de serre dans l’atmosphère tels que du dioxyde de carbone et du méthane. Pourtant, la composition de la communauté microbienne qui est à la base des processus biogéochimiques dans les mares de fonte a été très peu étudiée, particulièrement en ce qui concerne la diversité et l’activité des micro-organismes impliqués dans le cycle du méthane. L’objectif de cette thèse est donc d’étudier la diversité phylogénétique et fonctionnelle des micro-organismes dans les mares de fonte subarctiques en lien avec les caractéristiques de l’environnement et les émissions de méthane. Pour ce faire, une dizaine de mares ont été échantillonnées dans quatre vallées situées à travers un gradient de fonte du pergélisol, et disposant de différentes propriétés physico-chimiques. Selon les vallées, les mares peuvent être issues de la fonte de palses (buttes de tourbe, à dominance organique) ou de lithalses (buttes de sol à dominance minérale) ce qui influence la nature du carbone organique disponible pour la reminéralisation microbienne. Durant l’été, les mares étaient fortement stratifiées; il y avait un fort gradient physico-chimique au sein de la colonne d’eau, avec une couche d’eau supérieure oxique et une couche d’eau profonde pauvre en oxygène ou anoxique. Pour identifier les facteurs qui influencent les communautés microbiennes, des techniques de séquençage à haut débit ont été utilisées ciblant les transcrits des gènes de l’ARNr 16S et des gènes impliqués dans le cycle du méthane : mcrA pour la méthanogenèse et pmoA pour la méthanotrophie. Pour évaluer l’activité des micro-organismes, la concentration des transcrits des gènes fonctionnels a aussi été mesurée avec des PCR quantitatives (qPCR). Les résultats montrent une forte dominance de micro-organismes impliqués dans le cycle du méthane, c’est-à-dire des archées méthanogènes et des bactéries méthanotrophes. L’analyse du gène pmoA indique que les bactéries méthanotrophes n’étaient pas seulement actives à la surface, mais aussi dans le fond de la mare où les concentrations en oxygène étaient minimales; ce qui est inattendu compte tenu de leur besoin en oxygène pour consommer le méthane. En général, la composition des communautés microbiennes était principalement influencée par l’origine de la mare (palse ou lithalse), et moins par le gradient de dégradation du pergélisol. Des variables environnementales clefs comme le pH, le phosphore et le carbone organique dissous, contribuent à la distinction des communautés microbiennes entre les mares issues de palses ou de lithalses. Avec l’intensification des effets du réchauffement climatique, ces communautés microbiennes vont faire face à des changements de conditions qui risquent de modifier leur composition taxonomique, et leurs réponses aux changements seront probablement différentes selon le type de mares. De plus, dans le futur les conditions d’oxygénation au sein des
iv
mares seront soumises à des modifications majeures associées avec un changement dans la durée des périodes de fonte de glace et de stratification. Ce type de changement aura un impact sur l’équilibre entre la méthanogenèse et la méthanotrophie, et affectera ainsi les taux d’émissions de méthane. Cependant, les résultats obtenus dans cette thèse indiquent que les archées méthanogènes et les bactéries méthanotrophes peuvent développer des stratégies pour survivre et rester actives au-delà des limites de leurs conditions d’oxygène habituelles.
v
Abstract
The thawing and collapse of ice-rich permafrost in the subarctic region of Quebec has given rise to thaw ponds (thermokarst ponds) that emit the greenhouse gases carbon dioxide and methane to the atmosphere. However, the microbial community composition that underlies biogeochemical processes in thaw ponds has been little investigated, particularly concerning the diversity and activity of micro-organisms involved in the methane cycle. The objective of this thesis study was to determine the phylogenetic and functional diversity of micro-organisms in subarctic thaw ponds, and the relationships with environmental properties and methane emission. To that aim, we sampled ten thaw ponds in four different valleys located across a permafrost degradation gradient with distinct physico-chemical properties. Depending on valley, the ponds were derived either from the thawing of a palsa (peat-mound) or lithalsa (mineral-mound), which influenced the nature of organic carbon available for microbial remineralization. During summer, the ponds were observed to be well-stratified; there were with strong physico-chemical gradients down the water column, with an upper oxic layer and a bottom low oxygen or anoxic layer. To identify the factors influencing microbial community composition, we used high throughput sequencing techniques targeting transcripts of 16S rRNA gene, and additionally targeted genes involved in the methane cycle: mcrA for methanogenesis and pmoA for methanotrophy. As a proxy of microbial activity, we also measured the concentration of functional gene transcripts using with quantitative PCR (qPCR). The results showed a striking dominance of micro-organisms involved in the methane cycle, namely methanogenic Archaea and methanotrophic Bacteria. The pmoA analyses implied that methanotrophic Bacteria were not only active in the surface, but also in the bottom waters where oxygen concentrations were minimal; this was unexpected given their need for oxygen in methane consumption. In general, the microbial community properties were largely determined by the origin of the ponds (palsa versus lithalsa), and much less so by the extent of permafrost degradation. The key environmental variables pH, phosphorus and dissolved organic carbon likely contributed to the differentiation of microbial community between the palsa and lithalsa valleys. With intensification of climate warming, these microbial communities will face changing conditions that are likely to modify their taxonomic composition, and these responses are likely to differ between ponds in the two landscape types. Oxygenation of the ponds will likely be subject to major shifts in the future associated with changes in the duration of the ice-free season and the extent of stratification. Such changes will impact the balance between methanogenesis and methanotrophy, and thereby affect the net rates of methane emission. However, the results obtained here indicate that methanogenic
vi
Archaea and methanotrophic Bacteria have strategies to survive and remain active beyond the limit of their usual oxygen preferences.
vii
Table des matières
Résumé ... iii
Abstract ... v
Table des matières ... vii
Liste des tableaux ... x
Liste des figures ... xi
Liste des abréviations et des sigles ... xiii
Remerciements ... xvii
Avant-propos ... xviii
Chapitre 1. Introduction ... 1
1.1. Le pergélisol en Arctique ... 2
1.2. Les mares de fonte du pergélisol ... 3
1.2.1. Formation ... 4
1.2.2. Caractéristiques limnologiques ... 4
1.2.3. Implication dans les émissions de gaz à effet de serre ... 6
1.3. Les micro-organismes dans les milieux aquatiques ... 6
1.3.1. Processus microbiens ... 7
1.3.2. Micro-organismes impliqués dans le cycle du méthane ... 8
1.3.3. Approches pour étudier les micro-organismes ... 10
1.4. Organisation de la thèse ... 12
1.4.1. Objectifs et hypothèses des chapitres de thèse ... 12
1.4.2. Sites d’études... 13
Chapitre 2. Bacterial community structure across environmental gradients in permafrost thaw ponds: methanotroph-rich ecosystems ... 15
Résumé ... 15
Abstract ... 16
2.1. Introduction ... 17
2.2. Materials and methods ... 19
2.2.1. Study sites and sampling ... 19
2.2.2. Physico-chemical analysis ... 21
2.2.3. RNA collection and extraction ... 22
2.2.4. High throughput multiplex tag sequencing ... 23
2.2.5. Sequence processing and statistics ... 23
2.3. Results ... 24
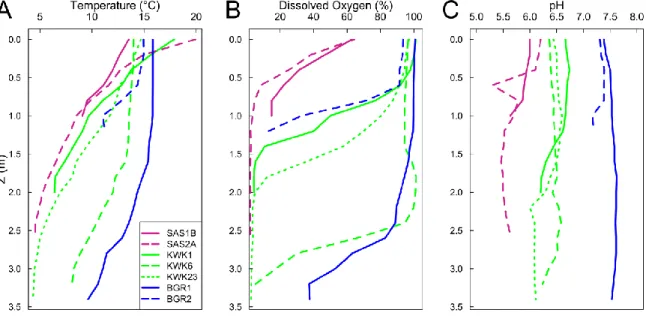
2.3.1. Limnological conditions ... 24
viii
2.3.3. Bacterial beta-diversity and community composition ... 26
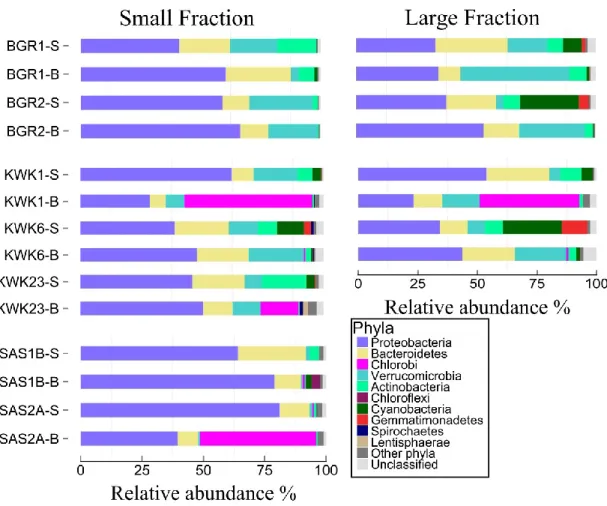
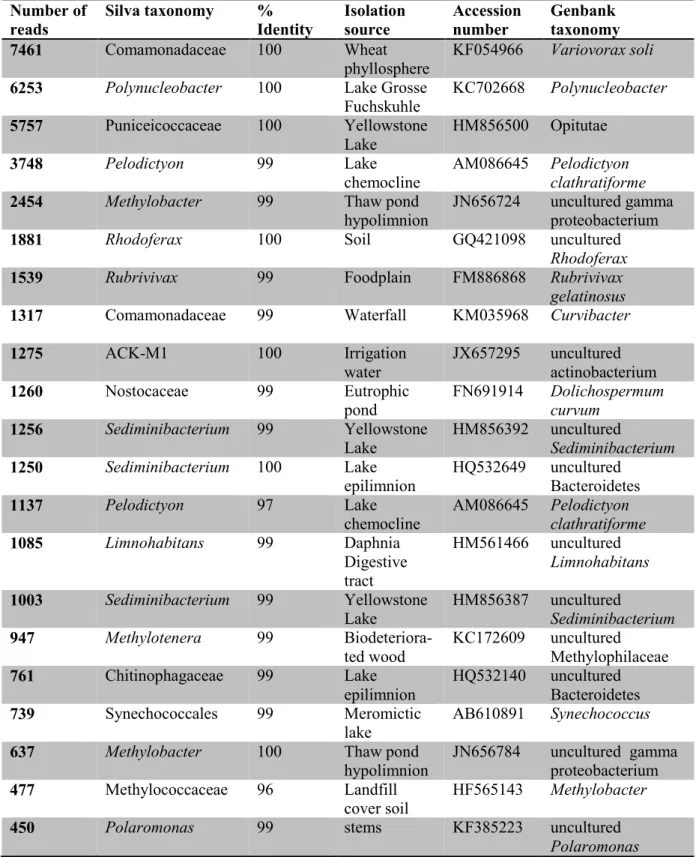
2.3.4. Bacterial dominants and high abundance of methanotrophs ... 27
2.3.5. Bacterial community structure as a function of environmental gradients ... 30
2.4. Discussion ... 32
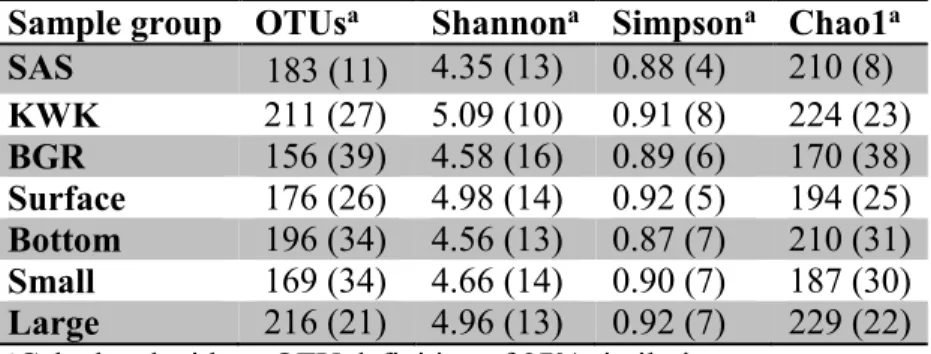
2.4.1. Bacterial alpha-diversity ... 32
2.4.2. Bacterial dominants ... 34
2.4.3. Attached and free-living Bacteria... 36
2.4.4. Depth gradients and bacterial community composition ... 36
2.4.5. Spatial variation and landscape gradients ... 37
2.5. Conclusions ... 38
2.6. Acknowledgements ... 39
Chapitre 3. Diversity and activity of methanotrophs in low-oxygen permafrost thaw ponds ... 40
Résumé ... 40
Abstract ... 41
3.1. Introduction ... 42
3.2. Materials and Methods ... 44
3.2.1. Study site and sampling ... 44
3.2.2. Sampling and physico-chemical measurements ... 44
3.2.3. RNA sample preparation and sequencing ... 44
3.2.4. Sample processing for qPCR ... 45
3.2.5. Sequence processing and analysis¸ ... 46
3.3. Results ... 47
3.3.1. Physico-chemical parameters ... 47
3.3.2. Community arrangement and composition ... 48
3.3.3. Methanotrophic activity ... 50
3.4. Discussion ... 53
3.4.1. Physico-chemical parameters ... 53
3.4.2. Community composition and arrangement ... 53
3.4.3. Methanotrophic activity ... 54
3.5. Acknowledgements ... 57
Chapitre 4. Environmental selection of planktonic methanogens in permafrost thaw ponds ... 58
Résumé ... 58 Abstract ... 59 4.1. Introduction ... 60 4.2. Results ... 62 4.2.1. Limnological conditions ... 62 4.2.2. Archaeal alpha-diversity ... 62
ix
4.2.3. Archaeal community dissimilarities and composition ... 64
4.2.4. Methanogens inferred from the mcrA versus 16S rRNA analyses ... 66
4.2.5. Environmental variables and archaeal community clustering ... 67
4.3. Discussion ... 68
4.4. Methods ... 71
4.4.1. Study site and sampling ... 71
4.4.2. Physico-chemical and molecular analysis ... 72
4.4.3. Bioinformatic analysis ... 72
4.5. Acknowledgements ... 74
Chapitre 5. Conclusion générale ... 75
5.1. Dominance des micro-organismes impliqués dans le cycle du méthane ... 75
5.2. Influence de l’origine des mares et du gradient de fonte du pergélisol ... 77
5.3. Influence des facteurs environnementaux ... 78
5.4. Implication scientifique ... 79
5.5. Perspectives ... 81
Bibliographie ... 84
x
Liste des tableaux
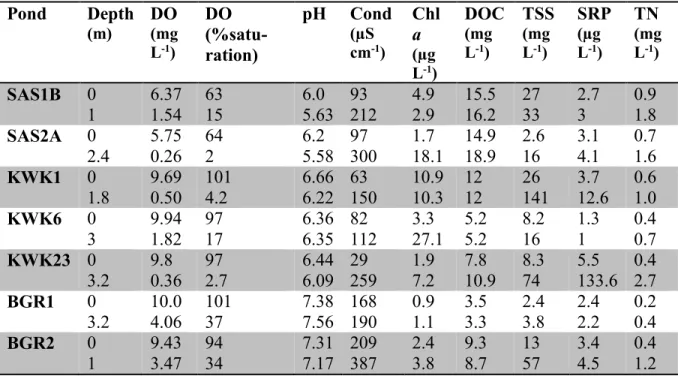
Table 2-1. Limnological properties of the sampled thaw ponds: dissolved oxygen (DO),
conductivity (Cond), chlorophyll a (Chl a), dissolved organic carbon (DOC), total suspended solids (TSS), soluble reactive phosphorus (SRP), and total nitrogen (TN). The surface samples correspond to 0 m and bottom samples to the second depth for each pond. ... 21 Table 2-2. Sequencing and diversity statistics for samples grouped according to valley, depth or size fraction. Values are means (n=3 to 14) with CV (SD as % mean) in parentheses. ... 26 Table 2-3. Identity of the 10 most abundant OTUs (defined at a level of 97% similarity) in each valley following the SILVA taxonomy. Following a BLASTn search, nearest matches and the providence of representative reads in GenBank were identified. Several groups appear multiple times because different OTUs match the same group. See Figure 2-5B for their distribution. ... 28 Table 3-1. Concentrations of pmoA transcripts oxygen and methane in the sampled thaw ponds. -: no data. ... 51 Table 4-1. Limnological properties of the bottom water (0.5 m above the sediment) for the sampled thaw ponds. Temperature (T°C), dissolved oxygen (O2), dissolved organic carbon (DOC), total
suspended solids (TSS) and total phosphorus (TP). ... 62 Table 4-2. Identity of the 5 most abundant OTUs (defined at a level of 97% similarity) for each valley following the lowest taxonomic level of the SILVA modified database (Lovejoy et al., 2015). The group Euryarchaeota, Miscellaneous Euryarchaeotic Group (MEG) and Thermoplasmatales could not be further assigned. Percent (%) represents the proportion of those single OTUs in the community for each valley. ... 66
xi
Liste des figures
Figure 1-1. Distribution du pergélisol en Arctique. Les deux nuances de violet plus foncées représentent le pergélisol de continu à discontinu tandis que les deux nuances de violet plus claires le pergélisol de sporadique à isolé (Brown et al., 1997) ... 3 Figure 1-2. Un exemple de paysage du pergélisol avec des mares de fonte, montrant la diversité des couleurs (site BGR, Nunavik; image : CEN/ArcticNet) ... 5 Figure 1-3. Processus microbiens tels que ayant lieu dans les mares de fontes du pergélisol. MOD désigne la matière organique dissoute. ... 8 Figure 1-4. Schéma simplifié des processus de méthanogénèse et méthanotrophie. ... 10 Figure 2-1. Location of the three sampling valleys in Nunavik, subarctic Québec, Canada. ... 20 Figure 2-2. Profiles of temperature (A), dissolved oxygen (B) and pH (C) as a function of the depth for the 7 studied ponds. ... 25 Figure 2-3. Relative abundance of the different phyla. The samples were from the surface (-S) and bottom (-B) of ponds in the three valleys. The small fraction (left) is for samples in the size range 0.2 to 3 µm, and the large fraction is for 3 to 20 µm. Phyla that were less than 1% of total
abundance are combined under “Other phyla”. ... 27 Figure 2-4. Relative abundance of methanotrophic Bacteria. The samples were from the surface (-S) and bottom (-B) of ponds in the three valleys. The small fraction (left) is for samples in the size range 0.2 to 3 µm, and the large fraction is for 3 to 20 µm. The taxa are labelled according to the highest taxonomical level; genera that represented less than 1% of total abundance were grouped together and labelled by their shared order. ... 29 Figure 2-5. (A) Bray-Curtis dissimilarity cluster analysis with the community data matrix (OTUs clustering at >97% of identity) for the study ponds. Surface samples are represented by triangles and bottom samples by circles, either filled (small fraction) or open (large fraction). (B) The bacterial dominants in each sample identified by their lowest taxonomical level either found on GenBank or SILVA (see Table 2-3); the size of the filled circle is proportional to their relative abundance. ... 30 Figure 2-6. Distance-based redundancy analysis ordination plot showing selected environmental variables that were significantly correlated with sample distribution. Abbreviations for the
environmental variables are given in Table 2-1. ... 32 Figure 3-1. Principal component analysis of the environmental variables: temperature (T), dissolved oxygen (O2), pH, total phosphorus (TP), total suspended solid (TSS), conductivity (Cond),
Chlorophyll a (Chla), total nitrogen (TN) and dissolved organic carbon (DOC) for the 9 sampled ponds. Colors indicate the different valleys; surface samples are represented by a triangle and bottom by a circle. ... 48 Figure 3-2. Bray-Curtis dissimilarity cluster analysis of the methanotrophic communities. Surface samples are represented by triangles and bottom samples by circles, either filled (small fraction) or open (large fraction). Heatmap shows the methanotrophic community composition. ... 49 Figure 3-3. Correlation plot of the PLS analysis to see influence of the environmental variables and methanotrophic genera on the activity of methanotrophs. Chosen environmental variables were conductivity (Cond), total phosphorus (TP), total suspended solid (TSS), Chlorophyll a (Chla), soluble reactive phosphorus (SRP), total nitrogen (TN), carbon dioxide concentration (CO2),
methane concentration (CH4), dissolved organic carbon (DOC), oxygen concentration (O2),
temperature (T°C) and pH. ... 52 Figure 3-4. Concentration of pmoA transcript as a function of methane concentration for bottom and surface samples (left panel), and for bottom only (right panel). Shading represents ± 95%
confidence intervals. ... 52 Figure 4-1. Alpha-diversity measures for the three sampled valleys. Shannon diversity index and Chao1 species richness index for the BGR, KWK and SAS valleys. The line in each box plot
xii
indicates the median, the box delimits the 25th and 75th percentile, and the whisker is the range. Diversity indices for SAS valley differed significantly from the two other valleys (p=0.02 for the Chao1 and p=0.01 for the Shannon index). ... 63 Figure 4-2. UniFrac clustering and composition of archaeal community. Upper dendrogram
representing the phylogenetic unweighted UniFrac distance of the archaeal community for the study ponds. Filled or open diamond represent respectively small and large fraction. First letter of sample name correspond to the valley name: S for SAS, K for KWK and B for BGR. The following number or combination of letter and number indicate the name of the pond. Bubble plot show the relative abundance of the different archaeal lineages with notably Miscellaneous Euryarchaeotic Group (MEG), Miscellaneous Crenarcheotic Groups (MCG) and Deep Euryarcheotic Sea Group (DESG). ... 64 Figure 4-3. Comparison of the relative abundance of the methanogens in the mcrA community and the 16S rRNA community. The left plot shows the entire methanogenic community and plot on the right shows the groups representing less than 1% of the reads. The SAS sample is the large fraction (3-20µm) of the SAS2B ponds and the KWK sample is the small fraction (0.2-3µm) of the KWK23 pond. ... 67 Figure 4-4. Non metric multidimensional scaling (NMDS) of the community composition.
Phylogenetic Unifrac distances are overlaid with environmental variables; (a) dissolved organic carbon, (b) total phosphorus and (c) pH. ... 68
xiii
Liste des abréviations et des sigles
16S : Petite sous unité ribosomique des bactéries et archées
ADN ou DNA : Acides désoxyribonucléiques ou Deoxyribonucleic acid ADNc ou cDNA : ADN complémentaire ou complementary DNA ANOVA : Analysis of variance
ARN ou RNA : Acides ribonucléiques ou Ribonucleic acid
ARNr ou rRNA : Acides ribonucléiques ribosomiques ou ribosomal RNA BGR : Bundesanstalt für Geowissenschaften und Rohstoffe
BLAST : Basic local alignment search tool BSA : Bovin serum albumin
CDOM : Coloured dissolved organic matter CEN : Centre d’études nordiques
CH4 : Méthane
Chl a : Chlorophylle a
CHUL : Centre hospitalier de l'Université Laval CO2 : Carbon dioxyde
Cond : Conductivity
dATPs : Deoxyadenosine triphosphate dbRDA : Distance-based redundancy analysis DMSO : Dimethylsulfoxide
dNTP : Deoxynucleotide triphosphate DO : Dissolved oxygen
DOC : Carbone organique dissous ou dissolved organic carbon DOM : Matière organique dissoute ou dissolved organic matter DSEG : Deep Sea Euryarchaeotic Group
GES : Gaz à effet de serre GF/F : Glass microfibre filter H2 : Dihydrogen
H2S : Hydrogen sulfide
H2SO4 : Sulfuric acid
HMM : Hidden Markov Models HSD : Honest significant difference
xiv KWK : Rivière Kwakwatanikapistikw
LBA : Luria-Bertaniagar
MCG : Miscellaneous Crenarchaeotic Group
mcrA : Sous-unité alpha de la méthyl coenzyme M reductase ou alpha subunit of the methyl- coenzyme M reductase
MEG : Miscellaneous Euryarchaeotic Group MES : Matière en suspension
mRNA : Messenger RNA NAS : Rivière Nastapoka
NCBI : National Center for Biotechnology Information NEB : New England Biolabs
NMDS : Non-metric multidimensional scaling nt : Nucleotides
O2 : Oxygen
OTU : Operational taxonomic units pb : Base pair (nucleotide)
PCA : Principal component analysis
PCR : Réaction en chaine par polymérase ou polymerase chain reaction pH : Potentiel hydrogène
PLFA : Acides gras dérivés des phospholipides ou Phospholipid-derived fatty acids PLS : Partial least squares
pMMO : Méthane mono-oxygénase soluble
pmoA : Sous-unité alpha de la méthane mono-oxygénase particulaire ou α subunit of the particulate methane mono-oxygenase
PVP : Polyvinylpyrrolidone
QIIME : Quantitative insights into microbial ecology qPCR : PCR quantitative
RDP : Ribosomal Database Project SAS : Rivière Sasapimakwananisikw
SIP : Marquage par isotopes stable ou stable-isotope probing sMMO : Méthane mono-oxygénase particulaire
SOC : Super Optimal broth with catabolite repression SRP : Soluble reactive phosphorus
xv TP : Total phosphorus
TSS : Total suspended solid
UTO : Unité taxonomique opérationnelle
V6-V8 : Région hypervariable de l'ARN ribosomique 16S ou hypervariable region of 16S rRNA gene
xvi
xvii
Remerciements
Tout d’abord, je tiens à remercier chaleureusement mon directeur Warwick Vincent et ma codirectrice de thèse Connie Lovejoy. Tout au long du parcours à embûches que constitue le doctorat, ils m’ont toujours montré un soutien sans faille et aussi une confiance en mon travail qui dépassait souvent la mienne. J’ai particulièrement apprécié la sagesse, la patience et la gentillesse de Warwick, qui a toujours vu le côté positif dans toutes les situations, ainsi que le franc-parler, l’humour et l’attention bienveillante de Connie. Merci à Steve Charette et Jean-Éric Tremblay pour leurs commentaires très constructifs durant les réunions de comité. Merci aussi à Philippe Constant d’avoir accepté de faire partie de mon jury. Ensuite je voudrais adresser un merci spécial à Jérôme Comte, qui a été très présent et disponible tout au long de ma thèse. Ses critiques et conseils toujours justes m’ont grandement aidée à bâtir une confiance en moi et une maturité scientifique. Merci à Marianne Potvin pour son aide précieuse au laboratoire et la mise en place de protocoles adaptés à mes échantillons. Ce doctorat n’aura pas été le même sans le temps passé avec tous les formidables collègues des labos Vincent et Lovejoy. Dans chacun d’eux, j’ai trouvé des qualités qui m’ont et continueront de m’inspirer pour devenir une meilleure scientifique. Merci à Paschale, Bethany et Mary dont l’amitié représente beaucoup pour moi en plus des dîners partagés ensemble et discussions motivantes. Enfin, j’ai eu la grande chance de partager un bureau avec deux personnes remarquables : Nathalie et Deo, avec qui l’entraide, le soutien, les débats et discussions stimulantes ont fait partie du quotidien. Une passion commune pour les soirées karaoké a contribué à établir un lien fort entre nous dont je serai toujours nostalgique.
Un tout grand merci chargé d’affection à ma famille, Papa, Maman, Isa, Ol, Fred, Julie et les petits bouts Manon et William pour leur soutien et amour inconditionnel. Merci à mes parents de toujours me soutenir dans mes choix, même si ça implique le déchirement de vivre éloignés. Merci à ma Maman et à Antoine d’avoir pris le temps de relire les parties de ma thèse en français. Merci à ma grande sœur et mon grand frère d’avoir été de si bons modèles dans ma vie de m’avoir toujours encouragé en plus de me donner de précieux conseils sur la vie universitaire.
Merci à tous mes amis de la Belgique au Canada. Sachez que tous les moments passés ensemble à s’amuser et à refaire le monde sont autant d’encouragements qui m’ont aidée à recharger mes batteries pour garder la motivation de continuer mon travail jusqu’au bout. Enfin, je remercie Antoine de remplir chaque jour de ma vie avec de la joie et des rires. Si j’ai pu mener à bien cette thèse comme je le voulais, c’est parce que j’ai toujours pu compter sur toi, sur tes conseils et quoiqu’il arrive, tu as inlassablement continué à me soutenir et à croire en moi.
xviii
Avant-propos
Cette thèse de doctorat dans le programme de biologie est présentée en cinq parties principales. Tout d’abord, une introduction explique le concept de mare de fonte du pergélisol et l’importance des micro-organismes impliqués dans ces milieux aquatiques, en particulier dans le cycle du méthane. Ensuite, les trois chapitres qui constituent le corps de la thèse sont des articles scientifiques rédigés en anglais dans le but d’être publiés dans des revues scientifiques révisées par des paires. Enfin, une conclusion générale résume et explore les implications des résultats obtenus pour les trois articles scientifiques. Les échantillons traités dans cette thèse ont tous été récoltés dans des mares de fonte du pergélisol dans la région subarctique du Québec dans le but de caractériser les communautés microbiennes de ces systèmes. Un intérêt particulier a été porté aux micro-organismes impliqués dans le cycle du méthane afin de mieux cerner leur diversité et activité. Pour chaque chapitre de thèse, j’ai contribué à la conception et la planification de la problématique ciblée avec l’aide de Warwick Vincent et de Connie Lovejoy, j’ai effectué la prise d’échantillons sur le terrain avec l’aide de Warwick Vincent, de Jérôme Comte et d’Alex Matveev, j’ai procédé à l’analyse en laboratoire et l’analyse bio-informatique avec l’aide de Jérôme Comte et de Connie Lovejoy et enfin j’ai rédigé tous les chapitres de thèse en tenant compte des contributions des coauteurs au niveau de l’apport de l’expertise scientifique, des corrections et commentaires. Les détails des articles scientifiques destinés à la publication se trouvent ci-dessous :
Chapitre 2. Bacterial community structure across environmental gradients in permafrost thaw ponds: methanotroph-rich ecosystems
Auteurs : Sophie Crevecoeur, Warwick F. Vincent, Jérôme Comte et Connie Lovejoy. Publié le 18 mars 2015 dans Frontiers in Microbiology, 2015.
Chapitre 3. Diversity and activity of methanotrophs in low-oxygen permafrost thaw ponds
Auteurs : Sophie Crevecoeur, Warwick F. Vincent, Jérôme Comte, Alex Matveev et Connie Lovejoy
En préparation, destiné à être soumis à un journal scientifique.
xix
Auteurs : Sophie Crevecoeur, Warwick F. Vincent et Connie Lovejoy
Soumis au journal Scientific Reports le 12 février 2016, présentement sous évaluation.
Les résultats de cette thèse ont également été présentés pendant plusieurs conférences internationales telles que la rencontre "Joint Aquatic Science Meeting" qui inclut la rencontre annuelle de l’"Association for the Sciences of Limnology and Oceanography" (ASLO), la rencontre ArcticNet "Arctic Change" et la conférence Embo "Aquatic Microbial Ecology SAME 14". De plus, des présentations orales ont été effectuées pour présenter les résultats de cette thèse dans plusieurs colloques prenant place à l’Université Laval comme lors du 81e congrès de l’association francophone pour le savoir (ACFAS), du colloque annuel de l’institut Hydro-Québec en environnement, développement et société et du colloque du département de biologie. J’ai personnellement conçu et présenté chacun de ces exposés oraux et et affiches dont les détails sont donnés ci-dessous, en tenant compte de la contribution des coauteurs.
Crevecoeur, S., Vincent, W. F., Lovejoy, C. 2015. Methanotrophic Bacteria in permafrost thaw ponds: potential for reduction of net methane emissions. Affiche présentée à la conférence Embo “Aquatic Microbial Ecology SAME 14”, Uppsala, Suède.
Crevecoeur, S. 2015. Détection des méthanogènes et méthanotrophes dans les mares de fonte du pergélisol. Présentation au colloque de biologie de l’Université Laval, Québec.
Crevecoeur, S. 2015. Biodiversité du Grand Nord en pleine mutation. Présentation au colloque annuel de l’institut Hydro-Québec en environnement, développement et société, Université Laval, Québec.
Crevecoeur, S., Vincent, W. F., Comte, J., Lovejoy, C. 2014. Molecular detection of methanogens and methanotrophs in permafrost thaw ponds: implication for greenhouse gas emissions from subarctic waters. Affiche présentée à la conférence ArcticNet “Arctic Change”, Ottawa.
Crevecoeur, S., Comte, J., Lovejoy, C., Vincent, W.F. 2014. RNA analysis of Bacteria in permafrost thaw lakes: implications for greenhouse gas emission. Affiche présentée à la conférence ASLO “Joint Aquatic Science Meeting”, Portland, Oregon, USA.
xx
Crevecoeur, S., Comte, J., Lovejoy, C., Vincent, W. F. 2013. Impact de la fonte du pergélisol dans la région subarctique du Québec et micro-organismes impliqués. Présentation au 81e congrès de l’association francophone pour le savoir (ACFAS) et symposium du centre d’étude nordique, Université Laval, Québec.
1
Chapitre 1. Introduction
Le réchauffement climatique est devenu un des enjeux majeurs de notre époque en raison de ses impacts et de son intensification. En effet, les émissions anthropogéniques de gaz à effet de serre venant des combustibles fossiles n’ont cessé d’augmenter depuis les années 1900 (IPCC, 2013). Parmi les conséquences du réchauffement global, on note une accumulation de chaleur dans les océans (Riser et al., 2016) et une augmentation anormale de la température des eaux profondes (Purkey et Johnson, 2010; Meehl et al., 2011), une forte diminution du couvert de glace notamment dans l’océan Arctique (Krishfield et al., 2014) et une fonte globale du pergélisol (Grosse et al., 2016). Ces changements risquent de s’amplifier dans le futur car les modèles développés pour l’instant prévoient des augmentations de température allant de 1 à 4°C d’ici 2100 (IPCC, 2013).
Les effets du réchauffement climatique sont en réalité plus intenses dans les régions nordiques car même de faibles augmentations de température peuvent avoir un impact important sur la fonte de la neige, de la glace et du pergélisol. En plus, une augmentation globale de la température de 2°C, qui représente un scénario moyen, correspondrait à une augmentation d’environ 8°C dans le Nord à cause notamment de différents effets de rétroaction positive comme la réduction de l’albédo, l’augmentation de l’humidité dans l’air (IPCC, 2013) et la libération de gaz à effet de serre provenant de la fonte du pergélisol (Vincent et al., 2013).
Ces effets présents et futurs ont attiré l’attention des scientifiques sur les lacs situés dans les régions nordiques car ces derniers constituent une des caractéristiques majeures du paysage (Grosse et al., 2013). Il y a de plus en plus d’évidences que ces lacs sont le théâtre d’activités biogéochimiques intenses, en particulier en ce qui concerne le cycle du méthane, un gaz avec un potentiel d’effet de serre environ 23 fois plus élevé que le dioxyde de carbone (Bastviken, 2009). Les lacs et mares arctiques sont parmi les principales sources de libération de méthane dans l’atmosphère (Wik et al., 2016). En effet, la fonte du pergélisol qui constitue le bassin versant des lacs nordiques résulte en un plus fort apport de matières organiques qui augmente l’activité microbienne à l’origine de la synthèse de gaz à effet de serre (Vincent et al., 2013). D’ici la fin du 21ème siècle, et en supposant une augmentation de la période libre de glace, on estime que les
émissions de gaz à effet de serre depuis les lacs et mares nordiques augmenteront de 20 à 54% (Wik et al., 2016). Plus particulièrement, les mares qui sont formées par la fonte, l’érosion et l’effondrement du pergélisol riche en glace (processus thermokarstique), représentent un type de milieu aquatique très biogéochimiquement actif.
2
Cette thèse porte sur l'étude de la composition de la communauté microbienne dans les mares de thermokarst (mares de fonte) en lien avec les processus biogéochimiques de l'environnement, plus particulièrement les micro-organismes impliqués dans le cycle du méthane. Ce chapitre d'introduction commence par la répartition du pergélisol en Arctique et explique les différentes caractéristiques de mares de fonte. Ensuite, plus de détails sont donnés à propos du rôle des micro-organismes dans les milieux aquatiques, que ce soit le recyclage de la matière organique ou leur implication dans les cycles biogéochimiques. Enfin, l'introduction se termine avec l'organisation de la thèse et la description des sites à l'étude. Cette recherche a eu lieu dans la zone subarctique du Québec, où une forte augmentation du réchauffement a été observée depuis les vingt dernières années, et où on constate des changements majeurs dans le paysage, incluant la dégradation du pergélisol et l’expansion et l’augmentation du nombre de mares de thermokarst (Bhiry et al., 2011).
1.1. Le pergélisol en Arctique
Le pergélisol, qui se définit comme un sol qui reste gelé (T°C ≤ 0) pendant au moins deux années consécutives (Dobinski, 2011), recouvre un quart des terres de l’hémisphère nord (Brown et al., 1998) (Figure 1-1). Le pergélisol contient aussi bien du sol riche en carbone comme de la tourbe ou du sol riche en minéraux (Vonk et al., 2015). Le pergélisol peut être continu (90-100% du territoire), discontinu (50-90%), sporadique (10-50%) ou isolé (2-10%) (Brown et al., 1998).
Au Canada, jusqu’à 50% du territoire contient du pergélisol (Smith et Riseborough, 2002). Dans le nord du Québec (région du Nunavik), tous les types de pergélisol sont présents et suivent un gradient, en partant du pergélisol continu au nord du 57e parallèle jusqu’au pergélisol sporadique
(défini comme moins de 2% du territoire par Allard et Seguin, 1987) au niveau du 54e parallèle
(Allard et Seguin, 1987). Cependant, avec l’intensification du réchauffement climatique, le pergélisol est soumis à une fonte estivale plus intense et la limite Sud du pergélisol remonte vers le Nord d’année en année (Halsey et al., 1995).
3
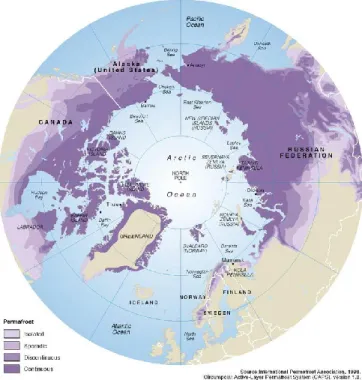
Figure 1-1. Distribution du pergélisol en Arctique. Les deux nuances de violet plus foncées représentent le pergélisol de continu à discontinu tandis que les deux nuances de violet plus claires le pergélisol de sporadique à isolé (Brown et al., 1997)
L’étude de l’impact de la fonte du pergélisol est un enjeu crucial puisque ce milieu constitue une réserve de carbone globale. En effet, la quantité de carbone organique stockée dans le pergélisol est pour l’instant deux fois supérieure à celle contenue dans l’atmosphère sous forme de dioxyde de carbone (Tarnocai et al., 2009). La fonte du pergélisol entraînerait donc une augmentation considérable de la concentration des gaz à effet de serre dans l’atmosphère avec comme conséquence une intensification accablante des effets du réchauffement climatique.
1.2. Les mares de fonte du pergélisol
Durant l’été, on peut constater l’apparition de mares de fonte de pergélisol (aussi appelées mares de thermokarst, voir Jansson et Taş, 2014). Au Canada, ces mares sont si nombreuses qu’elles constituent le type d’écosystème aquatique le plus répandu dans ses régions arctiques et subarctiques (Pienitz et al., 2008). En général, les mares de fonte rapetissent et/ou finissent par être drainées peu de temps après leur formation à cause de l’évaporation et de la dégradation du pergélisol (Smith et al. 2005; Riordan et al. 2006; Sannel et Kuhry 2011; Andresen et Lougheed 2015). Cependant, la région subarctique du Québec fait exception, car les mares de fonte persistent d’une année à l’autre (Smith et al., 2005). Ce phénomène s’explique par la présence de limon imperméable dans le sol, ancien bassin de la mer de Tyrrell présente il y a 8000 ans (Bhiry et al., 2011).
4
1.2.1. Formation
Dans les régions Arctiques où le pergélisol est continu, les mares de fonte se développent dans le centre de polygones et des ruisseaux se forment dans les coins de glace (Breton et al. 2009). Dans les régions subarctiques par contre, les mares sont issues de buttes de pergélisol (Calmels et al. 2008). Ces buttes sont appelées palses ou lithalses selon la composition de leur sol, respectivement à dominance organique ou minérale (Bhiry et Robert 2006; Calmels et al. 2008). De manière générale, les palses sont plus abondantes dans le pergélisol discontinu que les lithalses car les conditions climatiques propices à la formation de lithalses sont beaucoup plus restreintes par rapport aux palses. En effet, des conditions plus froides sont requises pour la formation de lithalses contrairement aux palses. Dans le nord du Québec, les conditions plus froides le long de la baie d’Hudson sont propices à la formation de lithalses alors que les conditions plus chaudes à l’intérieur des terres sont plus propices à la formation de palses, si bien que la majorité des zones de pergélisol discontinu dans le nord du Québec est dominée par des palses (Pissart 2000; Pissart 2002).
1.2.2. Caractéristiques limnologiques
Dans le nord du Québec, le gradient de fonte du pergélisol influence les caractéristiques du paysage, la géomorphologie des mares (Bouchard et al., 2014) et crée un gradient de concentration de la quantité de carbone allochtone contenue dans les mares. En effet, certaines mares de fonte sont entourées de pergélisol complètement dégradé alors que pour d’autres, environ 50% du pergélisol environnant est encore gelé. Ce phénomène crée une variation géographique des propriétés limnologiques des mares le long du gradient de fonte du pergélisol.
À l’échelle d’une vallée, des mares situées à quelques mètres les unes des autres présentent une grande variabilité de couleurs, reflétant leur diversité limnologique (Figure 1-2). L’étude de Watanabe et al. (2011) a montré que ces variations étaient dues en majeure partie au rapport entre les concentrations de carbone organique dissous, qui vient du pergélisol, et de matière en suspension, qui vient du sol environnant souvent riche en argile et limon, c’est-à-dire des particules en suspension qui ne sont pas d’origine algale. En outre, la concentration en nutriments et en chlorophylle a peut être très variable d’une mare à l’autre (Rossi et al. 2013). En général, les mares de fonte contiennent des concentrations élevées en nutriments allant de 6.2 à 91 µg L-1 pour le
phosphore ce qui correspond aux statuts mésotrophes et eutrophes (Brown et Simpson, 2001). Par contre, les données de chlorophylle indiquent souvent une faible productivité primaire à cause du contenu élevé en matière en suspension (MES) qui cause une grande turbidité et diminue donc l’efficacité de la photosynthèse. Néanmoins, l’abondance du bactérioplancton est assez élevée pour correspondre à des données propres aux lacs eutrophes (Breton et al., 2009; Laurion et al., 2010).
5
Figure 1-2. Un exemple de paysage du pergélisol avec des mares de fonte, montrant la diversité des couleurs (site BGR, Nunavik; image : CEN/ArcticNet)
Enfin, durant l’été, les MES absorbent les rayons du soleil ce qui réchauffe la couche supérieure de la colonne d’eau (épilimnion), créant une stratification thermique qui laisse la couche d’eau de fond (hypolimnion) plus froide. Même si les mares sont très peu profondes (entre 1 et 3 mètres de profondeur), la stratification reste très stable durant l’été et il n’y a pas de mélange entre l’épilimnion et l’hypolimnion (Laurion et al., 2010). Le brassage de la colonne d’eau a seulement lieu au printemps et à l’automne. Ces périodes de brassage peuvent même parfois être incomplètes et laisser l’hypolimnion isolé des couches d’eaux supérieures (Laurion et al. 2010; Deshpande et al. 2015). Cette forte stratification crée un gradient physico-chimique pendant l’été, car l’hypolimnion devient hypoxique à anoxique dû à la respiration bactérienne qui consomme presque tout l’oxygène présent dans la colonne d’eau et aucun brassage ne vient oxygéner l’hypolimnion isolé de l’atmosphère. Ce gradient physico-chimique a des conséquences pour les formes de vie au sein de la mare, car il crée des conditions propices au métabolisme anaérobie.
En résumé, les mares de fonte dans la région subarctique du Québec constituent un environnement hautement variable à différents niveaux, ce qui, d’un point de vue biologique, en fait
6
un système de choix pour étudier la biogéographie ou l’effet d’un gradient physico-chimique sur la composition des communautés planctoniques.
1.2.3. Implication dans les émissions de gaz à effet de serre
Depuis quelques années, l’attention des scientifiques pour les écosystèmes de mare de fonte s’est accrue en raison de leur contribution aux émissions de gaz à effet de serre (GES). En effet, le carbone resté prisonnier dans le sol pendant plusieurs centaines d’années devient alors disponible, avec la fonte, pour la reminéralisation microbienne. Les émanations de GES depuis les mares de fonte ne sont habituellement pas incluses dans les bilans globaux de budget de carbone puisque leur petite taille les rend invisibles aux outils de télédétection (Tranvik et al., 2009). Pourtant, leur contribution au cycle du carbone est considérable, surtout en ce qui concerne le méthane (Walter et al., 2007; Sepulveda-Jauregui et al., 2015) qui a environ 23 fois plus de potentiel en terme d’effet de serre que le dioxyde de carbone (Bastviken, 2009). Une estimation récente évalue la contribution des lacs et mares nordiques (au-delà du 50e parallèle) à environ 16 Tg de méthane par année (Wik et
al., 2016). Avec l’effet de l’accélération de la fonte du pergélisol, cette contribution peut augmenter considérablement. En effet, une étude de Walter et al. (2007) a estimé que la fonte totale du pergélisol sibérien entraînerait l’émission de 49 000 Tg de méthane dans l’atmosphère. Les émissions de GES depuis les mares de fonte constituent donc une rétroaction positive au réchauffement climatique (Walter et al. 2006).
Les mares de fonte dans la région subarctique du Québec ont des taux d’émission de GES très variables (Laurion et al., 2010). De manière générale, le dioxyde de carbone et le méthane s’accumulent dans le fond des mares avec des concentrations respectivement 8 à 16 fois et 2 à 125 fois plus élevées dans l’hypolimnion qu’à la surface (Breton et al. 2009). Le méthane peut s’échapper dans l’atmosphère par diffusion ou ébullition. Dans les mares de fonte situées en Alaska et en Sibérie, le processus d’ébullition est dominant pour le relargage de méthane dans l’atmosphère (Walter et al., 2008) et le flux diffusif ne représente que 6% des émissions de méthane de mares de fonte situées en Sibérie (Walter et al., 2006; Par contre, dans les mares de fonte située dans le nord du Québec, le méthane est émis principalement par diffusion (Matveev et al., 2016).Enfin, le mélange saisonnier permet la libération d’une grande partie du méthane accumulé dans l’hypolimnion (Bastviken, 2009).
1.3. Les micro-organismes dans les milieux aquatiques
Les micro-organismes dans les milieux aquatiques sont à la base des processus biogéochimiques de l’environnement. Dans cette thèse, la diversité des bactéries et des archées a été
7
étudiée. Ces deux domaines de la vie, malgré leur morphologie similaire, sont phylogénétiquement et physiologiquement distincts (Woese, 1996). Bien que l’écologie bactérienne soit connue depuis des siècles, les archées ont seulement été classifiées comme un domaine à part entière en 1990 (Woese et al. 1990). À cette époque, la connaissance de leur écologie se limitait aux environnements extrêmes. Cependant, les travaux de Delong (1998) notamment ont montré l’existence d’archées non extrémophiles. Il est maintenant reconnu que les archées sont présentes dans une grande variété d’environnements (Auguet et al., 2010) mais restent néanmoins plus difficiles à étudier en raison de leur faible abondance, comme c’est le cas dans les milieux aquatiques (Sievert et al., 2000; Chan et al., 2005).
1.3.1. Processus microbiens
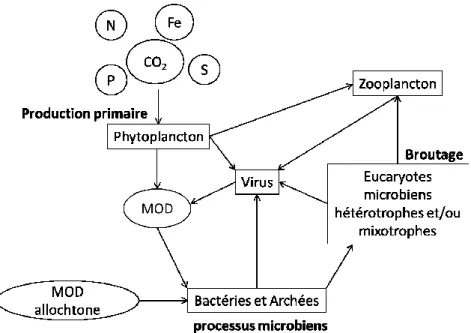
Un des rôles majeurs du plancton microbien dans les écosystèmes d’eau douce est le recyclage de la matière organique dissoute. Ce procédé est appelé « boucle microbienne », terme utilisé pour la première fois par Azam et Graf (1983) pour caractériser le rôle des microbes dans l’océan. Ce terme est maintenant largement utilisé pour les milieux d’eau douce également. Le principe de ce processus microbien est que les microbes planctoniques (bactéries et archées) consomment la matière organique dissoute autochtone (excrétion par le phytoplancton ou produit de la lyse virale) ainsi que la matière organique allochtone venant du bassin versant (figure 1-3). Au lieu de sédimenter, cette matière organique se trouve ainsi réintroduite dans la chaîne trophique par le biais du broutage des bactéries par les eucaryotes microbiens hétérotrophes ou mixotrophes. Les microbes ont donc un rôle prépondérant dans le recyclage de la matière organique dans les milieux aquatiques (Cotner et Biddanda, 2002; Fenchel, 2008). Les produits finaux de dégradation de la matière organique amènent à la formation de dioxyde de carbone dans des eaux oxiques, mais peuvent également mener à la synthèse de méthane en milieu anoxique.
8
Figure 1-3. Processus microbiens tels que ayant lieu dans les mares de fontes du pergélisol. MOD désigne la matière organique dissoute.
1.3.2. Micro-organismes impliqués dans le cycle du méthane
Environ 75% du méthane contenu dans l’atmosphère est d’origine biogénique (Chen et Prinn, 2005). Dans l’est de l’Arctique canadien, le méthane peut être d’origine moderne ou vieux de jusqu’à 4500 ans (Bouchard et al., 2015). Avec la fonte du pergélisol, une partie du méthane ancien resté prisonnier dans le sol gelé peut s’échapper dans l’atmosphère, tandis qu'une autre partie du méthane est moderne et nouvellement synthétisé par les archées méthanogènes, stimulées par la disponibilité en substrat venant de la fonte du sol. Les archées méthanogènes utilisent les substrats issus de la dégradation anaérobie de la matière organique comme l’acétate, les groupements méthyle, l’hydrogène et le dioxyde de carbone (Bapteste et al., 2005) pour synthétiser du méthane en milieu strictement anoxique (Thauer et al., 2008). L’utilisation de ces différents substrats détermine chez les archées l’utilisation de différentes voies métaboliques. En effet, les archées méthanogènes peuvent être acétotrophiques, méthylotrophiques ou hydrogénotrophiques selon les substrats qu’elles utilisent pour procéder à la méthanogenèse. On distingue sept ordres d’archées méthanogènes (Borrel et al., 2013) appartenant à deux lignées phylogénétiquement distinctes qui disposent d’un ancêtre commun, mais dont les descendants ne sont pas tous méthanogéniques (Bapteste et al., 2005). Cette distinction phylogénétique n’est pas liée à une différence de métabolisme, car tous les ordres de méthanogènes sont capables de procéder à la méthanogenèse hydrogénotrophique, mais seulement un ordre est capable d’utiliser les trois voies métaboliques (Bapteste et al., 2005).
9
En général, les archées méthanogènes peuvent être retrouvées dans différents types d’environnements, entre autres, les rizières (Wu et al., 2009), les tourbières (Yavitt et al., 2011), le pergélisol de la toundra (Barbier et al., 2012) ainsi que le pergélisol discontinu (Liebner et al., 2015). Dans les écosystèmes d’eau douce, les méthanogènes sont plus souvent associés avec les sédiments (Briée et al., 2007; Ye et al., 2009), comme c’est le cas pour les mares de fonte de l’est de l’Arctique canadien (Negandhi et al., 2013). La présence d’archées méthanogènes dans la colonne d’eau est moins sûre étant donnée la présence plus probable d’oxygène. Cependant, dû à la forte stratification et l’établissement d’un gradient physico-chimique, l’hypolimnion des mares de fonte peut disposer de conditions propices à la méthanogenèse en plus de fournir un apport riche en substrat venant de la fonte du sol (Laurion et al., 2010).
Une grande partie des flux diffusif de méthane vers l’atmosphère peut être oxydée par des bactéries méthanotrophes en entrant dans la couche oxique de la colonne d’eau. Ces bactéries ont la capacité d’utiliser le méthane comme seule source de carbone et d’énergie (Hanson et Hanson 1996). Les bactéries méthanotrophes sont historiquement connues comme appartenant aux branches alpha et gamma des protéobactéries (Bowman, 2006). Plus récemment, des nouveaux types de méthanotrophes ont été découverts, comme ceux appartenant au phylum des Verrucomicrobia qui ont la particularité de croître dans des conditions très acides (Dunfield et al., 2007) ou le phylum NC10 capable de coupler l’oxydation de méthane avec la dénitrification (Ettwig et al., 2009), se dispensant de ce fait du besoin d’oxygène comme accepteur d’électrons. Comme les méthanogènes, les méthanotrophes ne constituent donc pas un groupe monophylétique et l’apparition de ce trait semble s’être produite plusieurs fois au cours de l’histoire évolutive des bactéries.
Dans les lacs, les bactéries méthanotrophes peuvent consommer entre 30 et 99% du méthane (Bastviken et al., 2008). Par exemple, dans un lac boréal en Finlande, les méthanotrophes consommeraient 80% du méthane produit annuellement, qui se diffuse depuis les sédiments dans la colonne d’eau (Kankaala et al., 2006). Les méthanotrophes ont donc un rôle très important dans la régulation du climat en réduisant une forte proportion du méthane qui pourrait être émis dans l’atmosphère.
En résumé, l’équilibre entre l’activité des méthanogènes et des méthanotrophes détermine le bilan de carbone global et la quantité de méthane émise dans l’atmosphère. Cependant, des conditions très différentes sont requises pour la réalisation de l’un ou l’autre processus. Alors que l’oxygène inhibe la méthanogenèse (Bastviken, 2009), il est nécessaire à la méthanotrophie (Hanson et Hanson, 1996). Les mares de fonte ont la particularité de disposer d’environnements anoxiques et
10
oxiques au sein de la même colonne d’eau, les rendant particulièrement propices au développement des micro-organismes impliqués dans les processus de synthèse et de consommation du méthane (Figure 1-4).
Figure 1-4. Schéma simplifié des processus de méthanogénèse et méthanotrophie.
1.3.3. Approches pour étudier les micro-organismes
Depuis les 25 dernières années, le domaine de l’écologie microbienne a été révolutionné par le développement des techniques moléculaires (Dorigo et al., 2005; Pedrós-Alió, 2006). La première forme de technique de séquençage a été introduite par Sanger et al. (1977) et permet d’obtenir jusqu’à 1000 séquences d’un échantillon donné. Plus récemment, les techniques de séquençage à haut débit ont permis d’augmenter considérablement le nombre de séquences par échantillon. Par exemple, le pyroséquençage 454 permet d’obtenir de 1x105 à 1x106 séquences par
cycle et le séquençage illumina de 7x106 à 6x109 (Shokralla et al., 2012). Ces techniques passent
par la réalisation d’une PCR, ce qui peut occasionner des biais, mais nécessite aussi de cibler des gènes à amplifier pour répondre aux questions écologiques. Le gène le plus fréquemment utilisé pour étudier la diversité des bactéries et des archées est le gène 16S de l’ARN ribosomique (ARNr 16S) qui offre des résultats précis pour l’identification de la diversité microbienne avec les techniques de séquençage à haut débit (Liu et al., 2007). Cette approche, qui est adéquate pour étudier la diversité taxonomique des micro-organismes, ne suffit néanmoins pas pour évaluer leur diversité fonctionnelle. Pour se faire, il est nécessaire d’amplifier des gènes fonctionnels qui permettent de répondre à des questions concernant l’activité des micro-organismes. L’étude des gènes fonctionnels permet de relier la diversité microbienne associée aux processus biogéochimiques de l’environnement (Zak et al., 2006).
11
En ce qui concerne le cycle du méthane, plusieurs gènes ont été étudiés en lien avec les processus de synthèse et de consommation du méthane, ce qui facilite l’étude moléculaire des micro-organismes impliqués dans ce cycle. Au niveau du processus de synthèse, la méthyle coenzyme réductase catalyse l’étape finale de la formation du méthane (Ellermann et al., 1988) et le gène qui code pour cette enzyme (mcrA) est donc utilisé comme proxy pour étudier les archées méthanogènes (Luton et al., 2002). En ce qui concerne l’oxydation du méthane, l’enzyme méthane monooxygénase impliquée dans la première étape de cette réaction existe sous deux différentes formes : une enzyme particulaire (pMMO), qui est présente chez presque tous les méthanotrophes, et une enzyme soluble (sMMO) qui est seulement présente chez certains types de méthanotrophes (Murrell et al., 1998; Kolb et al., 2003). Le gène qui code pour la sous-unité alpha de la pMMO (pmoA) est plus particulièrement utilisé comme proxy pour étudier les bactéries méthanotrophes (Kolb et al., 2003). Cependant, à cause du caractère fortement paraphylétique de la méthanotrophie, il est impossible de disposer d’une séquence d’amorce permettant d’amplifier tous les types de pmoA (Sharp et al., 2012). La diversité des méthanotrophes au sein d’une même étude est donc souvent limitée à l’un ou l’autre groupe phylogénétique pour des raisons logistiques. Ces deux gènes marqueurs ont l’avantage de présenter une phylogénie cohérente avec celle obtenue grâce au gène 16S.
Au niveau moléculaire, il est possible de cibler l’ADN ou l’ARN du 16S et des gènes fonctionnels. Le choix d’utiliser une des deux ou les deux molécules dépend de la question de recherche. Que ce soit pour le 16S ou pour les gènes fonctionnels, le choix d’utiliser l’ADN permet d’étudier la composition de la communauté et la diversité des bactéries, des archées ou d’un groupe fonctionnel particulier. L’ADN permet de détecter non seulement les cellules vivantes, mais aussi les cellules mortes ainsi que des molécules libres d’ADN qui peuvent rester intactes dans des conditions froides ou anoxiques (voir Charvet et al., 2012). L’ARN permet aussi d’étudier la composition de la communauté et la diversité des micro-organismes, mais reflète par contre une communauté potentiellement active (Blazewicz et al., 2013), car les gènes sont à l’état de transcrits (Paul, 2001). L’étude de l’ARN requiert l’exécution d’une étape supplémentaire en laboratoire de transformation de l’ARN en ADN complémentaire (ADNc). En ce qui concerne les gènes fonctionnels, l’amplification de transcrits fournit une indication plus précise sur la présence de cellules métaboliquement actives (Juottonen et al., 2008). Bien que les techniques de séquençage évoquées au début de ce paragraphe permettent d’identifier une large portion de la communauté microbienne, ces approches restent semi-quantitatives. Pour mesurer le nombre de gènes ou de transcrits dans un échantillon, il faut procéder à une PCR quantitative (qPCR). Dans le cas de cette
12
thèse, la qPCR sur les transcrits de gènes fonctionnels a été utilisée comme technique pour évaluer quantitativement l’activité des groupes impliqués dans le cycle du méthane (Tuomivirta et al., 2009; Wilkins et al., 2015).
1.4. Organisation de la thèse
L’objectif général de cette thèse est de comprendre quels sont les facteurs qui influencent la composition de la communauté microbienne dans les mares de fonte de pergélisol ainsi que de connaître l’identité des micro-organismes impliqués dans le cycle du méthane. Les trois prochains chapitres, qui correspondent à des articles scientifiques, se concentrent sur la réalisation de ces objectifs.
1.4.1. Objectifs et hypothèses des chapitres de thèse
Chapitre 2 (article 1): "Bacterial community structure across environmental gradients in permafrost thaw ponds: methanotroph-rich ecosystems".
La forte stratification estivale crée un gradient de conditions physico-chimiques à travers la colonne d’eau des mares de fonte qui influence le métabolisme des micro-organismes et peut donner lieu à une séparation des communautés bactériennes. De plus, le gradient de fonte et de dégradation du pergélisol influence la quantité de matière organique allochtone présente dans les mares, ce qui a aussi un impact sur la composition de la communauté. L’objectif de ce chapitre est donc de comprendre les facteurs qui influencent la répartition de la communauté bactérienne et d’identifier plus particulièrement les bactéries méthanotrophes. Plus précisément les hypothèses suivantes ont été posées : (1) la communauté bactérienne qui se développe dans l’eau de surface des mares de fonte est différente de la communauté présente dans l’eau de fond à cause du fort gradient physico-chimique au sein de la colonne d’eau. (2) La composition de la communauté bactérienne varie en fonction du gradient de fonte et de dégradation du pergélisol. (3) Étant donné les fortes concentrations en méthane dans l’eau des mares de fonte, les bactéries méthanotrophes représentent en forte proportion de la communauté bactérienne.
Chapitre 3 (article 2): "Unexpected diversity and activity of methanotrophs in low-oxygen permafrost thaw ponds".
Le chapitre 3 a permis d’identifier une forte abondance de bactéries méthanotrophes à la surface et dans le fond des mares de fonte, mais les techniques utilisées dans le chapitre précédent ne permettent pas de savoir si ces bactéries participent activement à l’oxydation du méthane. Ce
13
chapitre-ci se concentre donc sur l’étude de la diversité fonctionnelle et l’activité des méthanotrophes pour mieux comprendre leur implication dans le cycle du méthane. L’objectif de ce chapitre est donc d’étudier la diversité fonctionnelle et l’activité des méthanotrophes ainsi que les facteurs qui les influencent comme l’origine de la mare ou le gradient de fonte du pergélisol. Ce chapitre se base sur les trois hypothèses suivantes : (1) la composition de la communauté des bactéries méthanotrophes dans les mares de fonte est déterminée par l’origine de la mare (fonte d’une palse ou lithalse) ainsi que par leur localisation au sein du gradient de fonte du pergélisol. (2) L’activité des méthanotrophes dépend de la composition de la communauté et est régulée par la disponibilité en oxygène au sein de la colonne d’eau. (3) Les bactéries méthanotrophes ont le potentiel de réduire les émissions nettes de méthane dans l’atmosphère.
Chapitre 4 (article 3): "Environmental selection of planktonic methanogens in permafrost thaw ponds".
Après avoir évalué la diversité et l’activité des méthanotrophes dans les chapitres précédents, ce chapitre se concentre sur les archées méthanogènes. Dans les écosystèmes lacustres, les conditions propices aux archées méthanogènes, c’est-à-dire l’absence d’oxygène, sont plus souvent retrouvées dans les sédiments que dans la colonne d’eau. Cependant, dans le cas des mares de fonte, l’eau du fond de la mare peut devenir hypoxique voir anoxique ce qui crée des conditions favorables pour la méthanogenèse. De plus, la fonte du pergélisol fournit un apport de substrat qui peut stimuler l’activité des méthanogènes. Ce chapitre a pour but d’étudier la diversité des méthanogènes planctoniques ainsi que les facteurs qui l’influencent comme l’origine de la mare ou le gradient de fonte du pergélisol. Plus particulièrement ce chapitre se base sur trois hypothèses : (1) l’eau de fond des mares de fonte constitue un habitat favorable pour les archées méthanogènes. (2) La composition de la communauté des archées dans les mares de fonte dépend de l’origine de la mare (fonte d’une palse ou d’une lithalse) et de leur localisation à travers le gradient de fonte du pergélisol. (3) Les mares issues de palses, qui sont plus riches en carbone, favorisent le développement d’une communauté d’archées plus diverse comparée aux mares issues de lithalse.
1.4.2. Sites d’études
Pour atteindre les objectifs et tester les hypothèses posées dans cette thèse, des mares de fonte situées dans quatre vallées différentes ont été échantillonnées. Ces vallées sont situées dans la région subarctique du Québec (Nunavik) à travers un gradient de fonte du pergélisol. Les deux vallées les plus au nord, se situent près du village inuit d’Umiujaq alors que les deux vallées les plus
14
au sud se trouvent près du village Kuujjuarapik où se côtoient les communautés Cris et Inuits. Plusieurs mares ont été échantillonnées dans chaque vallée en août 2012 et 2013.
1.4.2.1. Vallée de la rivière Kwakwatanikapistikw (KWK)
La vallée KWK se trouve à 12 km à l’est du village de Kuujjuarapik dans une zone de pergélisol sporadique (Bhiry et al., 2011). Dans cette vallée, la fonte des lithalses a donné lieu à la formation de mares de fonte qui recouvrent plus de la moitié de la surface du sol, témoignant de l’état avancé de dégradation du pergélisol (Bhiry et Robert, 2006).
1.4.2.2. Vallée de la rivière Sasapimakwananisikw (SAS)
La vallée SAS se trouve aussi dans la zone de pergélisol sporadique, à 8 km au sud de Kuujjuarapik. Cette vallée abrite une tourbière ombrotrophe, c’est-à-dire que l’eau qui l’alimente ne vient que des précipitations et est donc souvent acide et pauvre et nutriments. Dans cette vallée, le pergélisol est très dégradé et l’effondrement de palses donne naissance aux mares de fonte (Arlen-Pouliot et Bhiry, 2005).
1.4.2.3. Vallée “Bundesanstalt für Geowissenschaften und Rohstoffe” (BGR)
La vallée BGR se situe plus au nord à environ 20 km à l’est d’Umiujaq, dans une zone de pergélisol discontinu et répandu, c’est-à-dire que plus de 50% du territoire est composé de pergélisol. Cette vallée est recouverte de limon marin postglaciaire et de tourbe, par conséquent les mares y sont issues de palses et de lithalses (Calmels et Allard, 2004; Calmels et al., 2008). Dans le cadre de cette étude et pour des raisons logistiques, seules les mares issues de lithalses ont été échantillonnées dans cette vallée.
1.4.2.4. Vallée de la rivière Nastapoka (NAS)
La vallée NAS représente la vallée la plus au nord échantillonnée pour cette thèse. Elle se situe dans une zone de pergélisol discontinu et répandu à environ 30 km au nord d’Umiujaq. Les mares de fonte dans cette vallée ont encore été très peu étudiées. Les mares échantillonnées dans la vallée NAS dans le cadre de cette étude proviennent de la fonte de lithalses (Seguin et Allard, 1984).
15
Chapitre 2. Bacterial community structure across
environmental gradients in permafrost thaw ponds:
methanotroph-rich ecosystems
1Résumé
La fonte du pergélisol mène à la formation de mares de thermokarst qui émettent potentiellement du CO2 et du CH4 dans l’atmosphère. Dans la région subarctique du Nunavik (Nord
du Québec, Canada), ces nombreuses mares peu profondes deviennent fortement stratifiées pendant l’été. Cela crée un gradient physico-chimique de température et d’oxygène, avec une couche d’eau de surface oxique et une couche d’eau de fond pauvre en oxygène ou anoxique. Notre objectif était de déterminer l’influence de la stratification ainsi que des propriétés limnologiques et du paysage sur la structure des communautés bactériennes potentiellement actives dans ces eaux. Les échantillons pour l’analyse ARN ont été pris dans des mares situées dans trois vallées différentes à travers un gradient de fonte du pergélisol. Un total de 1296 unités taxonomiques opérationnelles a été identifié par séquençage d’amplicons à haut débit, ciblant l’ARNs 16S bactérien, préalablement retrotranscrit en ADNc. Les betaproteobactéries étaient le groupe dominant dans toutes les mares, avec la plus forte représentation par les genres Variovorax et Polynucleobacter. Les méthanotrophes étaient aussi parmi les séquences les plus abondantes pour la plupart des sites. Ils comptaient même pour 27% du total des séquences (médiane de 4.9% pour tous les échantillons), indiquant l’importance du méthane comme source d’énergie bactérienne dans ces eaux. Des phototrophes oxygéniques (Cyanobactéries) et anoxygéniques (Chlorobi) étaient aussi fortement représentés, ces derniers dans les eaux de fond pauvres en oxygène. Des analyses d’ordination ont montré que les communautés se regroupaient en fonction des vallées et de la profondeur, avec un effet significatif attribué à l’oxygène, au pH, au carbone organique dissous et aux matières en suspension. Ces résultats indiquent que les assemblages bactériens dans les mares de fonte du pergélisol sont filtrés par les gradients environnementaux, et représentent des associations complexes de taxons fonctionnellement divers qui ont de grandes chances d’affecter la composition ainsi que la magnitude des émissions de gaz à effet de serre depuis ces abondants plans d’eaux.
1 Citation : Crevecoeur S, Vincent WF, Comte J and Lovejoy C (2015). Bacterial community structure across environmental gradients in permafrost thaw ponds: methanotroph-rich ecosystems. Front. Microbiol. 6:192. doi: 10.3389/fmicb.2015.00192
16
Abstract
Permafrost thawing leads to the formation of thermokarst ponds that potentially emit CO2
and CH4 to the atmosphere. In the Nunavik subarctic region (northern Québec, Canada), these
numerous, shallow ponds become well-stratified during summer. This creates a physico-chemical gradient of temperature and oxygen, with an upper oxic layer and a bottom low oxygen or anoxic layer. Our objective was to determine the influence of stratification and related limnological and landscape properties on the community structure of potentially active Bacteria in these waters. Samples for RNA analysis were taken from ponds in three contrasting valleys across a gradient of permafrost degradation. A total of 1296 operational taxonomic units were identified by high throughput amplicon sequencing, targeting bacterial 16S rRNA that was reverse transcribed to cDNA. Βetaproteobacteria were the dominant group in all ponds, with highest representation by the genera Variovorax and Polynucleobacter. Methanotrophs were also among the most abundant sequences at most sites. They accounted for up to 27% of the total sequences (median of 4.9% for all samples), indicating the importance of methane as a bacterial energy source in these waters. Both oxygenic (cyanobacteria) and anoxygenic (Chlorobi) phototrophs were also well-represented, the latter in the low oxygen bottom waters. Ordination analyses showed that the communities clustered according to valley and depth, with significant effects attributed to dissolved oxygen, pH, dissolved organic carbon, and total suspended solids. These results indicate that the bacterial assemblages of permafrost thaw ponds are filtered by environmental gradients, and are complex consortia of functionally diverse taxa that likely affect the composition as well as magnitude of greenhouse gas emissions from these abundant waters.