/ .
"\
'
,,4
J ' Etude socio-êco1ogique de}!-l'Etang de
Mi~agôane,
Haïti.par
@Jean
Phaneuf..
Thèse soumise à la Faculté des Etudes Graduées et de la Recherche, à titre d'apport partiel en vue de l'obtention du grade d'e Maîtrise ès Sc iences .
Département des Ressources Renouv~lables
Université McGill /
ABREGE Tllès-e de
maîtrise
Jean Phaneuf Ressources Renouvelables
"ETUDE SOCIO-ECOLOGIQUE DE L'ETANG DE MIRAGOANE, HA rTI '!
En 1951, le cichlidé africai,n, Sarotherodon mossambiou8, fut, introduit pour pallier à la faible productivité ichthyologique des eaux douces d'Haïti. Du mois d'août 1978 au mois de février 1979
, nous analysons la population des cichlidés de l'Etang de Miragôane, en Haïti. Nous y évaluons le, succès d'implantation du cichlidé africain, sa relation avec l'espèce indigène, CiohZa8oma haitiensis et nous quantifions 1 '.utilisation des divers habitats lacustres. Nous étudions aussi l'apport économique de ces ressources et celui des "joncs", CZadiumjamaicensis-, Eleooharis sp.
L'introduction du cichlidé africain a grandement amélioré la productivité des poissons, sans pour autant engendrer de compétition
"
réelle avec le cichlidé haitien quoiqu'ils utilisent le même habitat ~,' pour la reproduction et l'élevage, soient les terrasses du littoral.
Les pêcheries n'utilisent qu'une infime partie des 111,~OO kg
de poissons produits annuellement et elles rapportent environ $12,OOO/an pour 170 pêcheurs. Par contre les marais d'éléocharides engendrent
$117,000/an répartis entre 631 femmes. Ains1, ces ressources
entraînent des revenus annuels de $130,000. Par contre ces intrants économiques pourraient être améliorés par des meilleures techniques de pêche et par l'utilisation annuelle de 2,000 rn-hectare d'eau
( pour irriguer 1500 hectares de la plaine adjacente.
ABSTRACT M.Sc.
thesis
Jean Phaneuf Renewable Resources
"SOCIO-ECOLOGICAL STUDY OF ETANG DE MlRAGOANE, HAITI'!
(
The african cichlid, SaPotherodon mossambicus was introduçed to the Etang de Miragôane~f Haiti in 1951 ta increase the fish produc-tivity. From August 1978 ta February 1979 we evaluated the success of the introduction, the relationship between the African and 'the endemic species of cichlid, Cichtasoma haitiensis~ and quantiEied
local use of the aquatic resources including the harvest of the emergent plants Ctadium jamaiaensis and Eteoaharis sp.
t
The introduction of S. m08sambiau8 has great1y improved the fish productivity of the Etang de Miragôane. The resu1ts indicate that there is 1ittle competition between the 2 species although bath are dependant upon the terraces of the littoral zone as spawning are as and nurseries.
The local fisheries provides employment for 170 male Haitians and yield approximately $12,000 annually ta the local economy, but
only a small proportion of the annual fish production of 111,000 kg/year i9 utilized. The aquatic' emergent spike rush i8 harvested by
approximate1y 631 females yielding a gross annuai input to the local economy of $117,000. Thus the aquatic resources of the Etang de
A~
Miragoane presently support over 800 families and provide a direct economic input of $130,000 per year.
!
1
- lt is suggested that this value may be increased by improving
local fishery techniques. Furthermore, 2000 rn-ha of water may be
used annually for the irrigation of a proposed 1500 ha of land
without any significant negative impact on the local aquatic resources.
/ '
')
,
-. REMERe IEMENT S
" .1
TABLE DES MATIERES
..
"',...
,... .
, ; l ' Page iv CONTRIBUTIONS ORIGINALES 1 • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • •• • • • • vi / 'LISTE DES TABLEAUX ••• :" •.•.•••••.••.•••.•••••.•.•••..••...•....•••
.
u
vii LISTE 'DES FIGURES ...•...•••...••...•... :-.•.. .'... viii , AVANT-PROPOS ••••.••••.••••.•••...•••••..•••••••.•••••.•••••....•• ,Chapitre 1. Les caractères biologiques et la compétition inter-spécifique des 2 cichlidés de l'Etang de Miragôane, Haïti;'
Cichtasoma haitiensis
Tee Van,Sal'otherodon
mossambicus
Pet ers •••••••...••••••••••••.••.••••••••1.1 INTRODUCTION .•••...••...•.•...•.•...••....•.••...••. 1.1.1
1.1. 2 1.1.3
Les antécédents .•••••.••••..•••.••..•••..•.••••..•••• Les obj ectifs ...•••...•. : ...•...•....•.•••....••
Le site .. III • • • • • • • • • • • • • • • • • • • • • • • • • • ~/. 'II • • • • • • • " . . . 1. 2 MATERIEL ET METHODES ...•••.••••••...••••.•.•••.•..••••••.••• \. 1.2.1 Le matériel . . . ~. ~ ... . 1.2.2 Les méthodes ••...••..••••••...••••.•.•••...•••••...•• 1.2.2.1 1.2.2.2 1.2.2.3 1.2.2.4
L'enregistrement des données ••.•.•••.•••..•• L'étude de la longueur •••.•.••...•••••••..•• L'étude de la masse ..••...•.••...•. :~ •.•..•• L'étude de l'embonpoi~t .•••.•••..•••••....••
"
1.3 RESULTATS: Les paramètres biologiques des cichlidés de
l'Etang de Miragôane •••.•.••••••••••..•••••••••••
1.3.1 Sarothel'odon mossambicus
Peters •.••..•••...••••....••1.3.1.1 La longueur de
S. mossambicus ... .
1.3.1.2 La masse de
S. mossambicu8 ....•...•
1.3.1.3 L'embonpoint de
S. mossambicus ... .
1.3.2 Cichtasoma haitiensis
Tee Van . . . ..1.3.2.1 1.3.2.2 1.3.2.3
La longueur de
C. haitiensis ... .
La masse de
C. hai tiensis ... .
L'embonpoint
de
c.
hàitiensis ...•...•
i 1 3 5 5 5 6 6 6 6 6 7 7 / 8 9-9 9 9 . 10 10 10 11 I l1. 3.3 Parallèles biologiques entre·tees cichlidés
1.3.3.1 La densité . . . • . . . 1.3.3.2 La longueur . . . • : ...••... 1.3.3.3 La masse . . . • • . . . 1.3.3.4 L f embonpoint . . . . 1.3.3.5 Des observations diverses ...•...
12 12 12 12 l3 13 1.4 DISCUSSION •...•••••...•••••.•.•••••.•.•••••••••••.•.•••••.•.• 13 1.4.1 Le succès de S. mossambicus ... 13
1.4.2 L'infériorité de C. haitiensis et la compétition avec S. mossambicus ....•...•...•.... 15
1. 5 CONCLUSION •.•.••.••.•.•••••...•••••...••••...••.•...••...• , 16
Chapitre 2. Analyse descriptive de l'Etang de Miragôane, Haïti, et répartitio~ des cichlidés, Cichlasoma hattiensis Tee Van et SaY'otherodon mossambicus Peters, en 2.1 fonction des milieux . . . • . . . . • . . . : ... -.~ ... . INTRODUCTION 2.1.1 2.1. 2 2.1. 3 Les ant~dents ..•.•...••...•....•..•...•••.... Les objectifs . . . • • . . . . • . . . • • • . . . • Le site . . . . 17 19 19 _/ 19 20 2. 2 MATERIEL ET :METHODES .•.•••••••.••••.•••••.••••••••••••••••• -; • 20 2.2.1 2.2.2 2.2.3 La géographie physique . . . • . . . • . . . L'analyse physico-chimique de l'eau . . . • . . . L'échantillonnage ichthyologique . . • . . . < • • • • • • • • • • • 20 21 21 2.3 RESULTATS .••...• : . • • . . . • • • . . . • • • • • . . . • • . . • • . • • . • . . . • • • . • . • . 22 2.3.1 L'Etang de Miragôane . . . 22
2.3.1.1 Les caractères physiques... 22
2.3.1.2 Les associations végétales . . . 23
2.3.1.3 Les milieux • . . . 24
2.3.2 La répartition des cichlidés en fonction des plans d'eau . . . , . . . 25
2. 3'~ 2.1 SaY'otheY'odon mossambious .•... 25
2.3.2.2 Cichlasoma haitiensis .... ... ... 26
2.3.3 La répartition des cichlidés en fonction des milieux.. 27
2.3.3.1 S. mossl1.lrbicus .•.•.•...•••...•.... 27
2.3.3.2 C. haitiensis ... _ ..
.,j'...
282.4 DISCUSSION 28
.
" 2.4.1 2.4.2;
S. mossa:nbicus ... : { .•...•....•....•...•.. C. hai tiensis ... / ... . 2.5 CONCLUSION .•••.••.•...•.• ./ ...•••.••.•••...•...•...••. 1 1Chapitre 3. L'utilisation actuel1~' des ressources de l'Etang de Miragôane, Haïti, dans la vie rurale, et
l'optimisa-3.1
tion possible de ce potentiel . . . .. INTRODUCT ION
3.1.1 3.1.2 3.1. 3
Les antécédents .•... : . . • . . . • . Les obj ec tif s ....•.••.••..•..•.•...•..•.••....•..•.•• Le site . . . . 3.2 MA.TERIEL ET METHODES .•..•...••..••••••..•.•..•.•.•..•...• 3.2.1 3.2.2 Les recensemente . . . • . . . • . . . • A h '1 i
Les techniques de peche a~t enne •.•..•.•. ~ ...•.••.•.
3.3 RESULTATS •.••••.•••...•....• , •....•....•.•.•.•••...•..••••
3.3.1 Les ressolirces halieutiques •..•...•.•....•..•. 3.3.1.1
3.3.1.2 3.3.1.3
- h h'"
Le pee eur al. t l.en . . . • . . . • . Le rendement des techniques halieutiques •.•• La "t1adame Sarah", •..•.•....•....••...•. 3.3.2 Les coupeurs de "joncs" . . . .
3.4 DISCUSSION .oi •••••••••••• ~ •••••••••••••••••••• f • • • • • • • • • • • • • • •
3.4.1 Les pêcheries: son utilisation actuelle et son
• potenl=iel réel ••••.•..••...•.•.•.•.••...••.•..•...
3.4.2 La végétation marécageuse ...••...•....•...•. 3.4.3 L'irrigation en période s~ehe •••.••.•....•..•...••
3 • 4 CONCL US ION •••.•.•••••.•.•..••••••...•••.•.••....•....•.•..•• SOMMAIRE ..•••••..•.•.••.•.•..•.•..•...••.•...••....•....•..•.•. / BIBLIOGRAPHIE ••..•••..••...••...•••..•.•.•..•.•....•....•....•. TABLEAUX •..••.••.•.••.•...•..•.••.•••..••.•••.•.••...•.•..•.••.•• FIGURES Hi Page 28 30 30 32 34 34
34
35 35 35 36 38 38 38 39 40 4244
44 46 46 48 49 50 53 60 ",
o REMERe IEMENT
S'
Je remercie mon directeur de thèse, le Dr. J.R. Bider, pour , les encouragemènts et les conseils prodigués tout au long de ce
travail.
, /
Je tiens à souligner le rôle essentiel de l'Agence Canadienne
de Développement International (A.C.D.I.) qui a assumé la logistique
de cette étude et qui m'a octroyé une bourse d'étude substantielle.
Relativemen~ à cet organisme je remercie particulièrement les
personnes, suivantes:
_ /
Le Dr. Gilles Blanchet de la Direction de l'Evaluation à
l'A.C.D.I. pour son support ~onstant.
M . ean-Yves Cote, a ors J A ~ 1 Ch arge ~ d e P rojet pour a1t1, pour H '1 •
son aide administrative.
l '
M. Edmond Magny, alors Directeur des 'Etudes au programme
1 ~
lJ
-D.R.I.P.P. en Ha1ti, pour son soutien technique.
M. Antoine Assaf, aloys Chef du Génie rural au programme
D.R.I.P.P., pour l'échange des données géophysiques, liées au
proj et de drainage de l'Etang de Miragôane. \.
Le Dr. Thai Cong-Tung, agronome-pédologue du programme
D.R.I.P.P., pour sa contribution à l'identification de
la
zone.Je remercie également le Dr. Albert Stauch/ conseiller de
la Food Agriculture Organization (F.A.O.) attaché au Service des
Pêcheries d 'Haïti, pour sa coopé~ation et son suivi lors de l'échantillonnage sur le terrain.
Je remercie finalement tous cêux, qui ont collaboré dè près ou de loin à cette étude et dont l'énumération complète serait trop longue. / () , 'i v • J
/
, ,
/
CONTRIBUTIONS ORIGINALES l '
1. Nous évaluons le succès de
S. moltambicus
en Amérique latine,, ,~
quelques 25 années après son introduction, par comparaison avec s9n homologue haÏtien,' C.
ha1-tiensis.
2. Nous déterminons la crbissance en poids du cichlidé haitien Ci::- J dans les eaux naturelles cl' Haïti.
-.
3. Nous déterminons la croissance en poids de S.
mossambicus
dans les eaux naturellesd'HaïtI:-4. !pus quantifions les migrations de S.
mossambicu8
et de C.haitiensis
parmi les habitats lacustres et, selon leur stade decroissance, une première en Hispaniola.
5. Nous évaluons l'efficacité des techniques de pêche haïtienne
su~
les populations de cichlidés en Haïti, par comparaisonavec la technique universelle des filets maillants expérimentaux. 6. Nous identfJions socialement l'usager haïtiên des ressources
aquatiques télles que les pois,~ons , et la végétation émergeante.
\
-.--/' 7. Nous déterminons l'apport économique en milieu rural haïtien
\
\
\
\
des pêcheries de cichlidés et des macrophytes émergeant du littoral. vi
\
\ \ \-=.
--..
-,> , ", ,(,
.
Tableau 1. / Tableau 3. Tableau 4.LISTE DES TABLEAUX
Composition de la population des cich.1idés de ' ~Etang de Miragôane ....•....•... :"':' . . . . Médianes 'des paramètres' de croissance selon les groupes d~ longu;ur, pour les cichlidés de l'Etang de Miragôane .~ . • . . . • . . . • . . . • . . .
\ ' /
Médianes des par~mètres biologiques des cich1idés de l'Et.ang de Miragôane en fonction de leur distri-bution . . . • • • . : ...•...•.... Répartition de
S. mossambiouB
en fonction des lacs de l'Etang de'.Miragôane ••..•..•.• , ..•.•••....•. , .•.. Tableau 5. Répartition de C. -Haitiensis en fonction des lacsde l'Etang de Mirag5ane . . . ' . . . . .. Tableau 6.
Tableau' 7.
1
-
,Répartition de
S: mossambiouB
en fonction desmili~ux de l'Etang---d'e Miragôane •...••...•••.•• Répartition de C. haitiensis en fonction des milieux de l'Etang de Miragôane ••. ' .•..•.•....•.•...•...•. " vii
o.
.,r Page,,'
53 i .. 54 " 55 56 0 57 58 59J
, 1o
•
/)
LISTE DES FIGURES
Figure 1. Histogrammes des fréquences de longueur pour les cichJidés de l'Etang de Miragôane •..••.••••.•••••. Figure 2. Relations Longueur-Masse des cichlidés de l'Etang de Miragôane et celles de S. mOBsambicu8 issues --de régions tropicales et néo-tropicales .•.••••..••••• Figure 3. Variations du coefficient de condition chez les
Page
60
62
cichlidés de l' Etang ~e Miragôane . " . . . • . . . • . . 64
Figure 4. L'Etang de Miragôane, Ha1ti, et la vocation des sols /'J '
adj acents "' . . . . 66
Figure 5. Profils phy~ico-chimiques de l'eau de l'Etang de
Miragôane, Haïti •••••...•....•.. ,', ...•...•.•. 68 Figure 6. Profil du littoral de l'Etang de Miragôane, ~ït1 70
J
viii
/ '
/
f
(
AVANT-PROPOS
- / L'intérêt de l'auteur pour l'Etang de Miragôane repose sur deux
prémisses: ce complexe lacustre constitue le deuxième en importance
en Haïti et une saine gestion de ses ressources s'avère non seulement
souhaitable, mais essentielle pour ce pays ~urpeuplé constamment à
. /
la recherche de ressources alimentaires. Le cichlidé africain
S. mossambiaus
Peters constitue plus de 70% des captures dans celac et l'aménagement de-cette zone marécageuse se doit de t~nir
compte des contraintes
écol~giques
de ce poisson afin de préserver cette source nutritive importante.~ien que la~gement implanté dans les eaux douces.tropicales
et néo-tropicales, l'évaluation du succès de l'introduction de
,
S. mossambiaus
et la connaissance de son écologie dans les eaux, / colonisées, demeurent très superficielles comparativement à
l'impor-tance du suj et.
Cette étude présente donc une appr~ciation de la population
des cichlidés de l'Etang de Miragôane et de l'utilisation des ,
ressources tirées de l'Etang de Miragôane par le paysan haïtien.
Quant au plan de travail même, le premier chapitre compare la
croissance et la densité des deux cichlidés,
S. mossambicus
etC. hairensis.
de ces poissons
La seconde section étudie la distribution biologique
parmi les habitats lacustres. Le dernier chapitre
expose les répercussions des ressources de l'Etang de Mirag6ane sur ,
l'économie rurale et discute de possibilités d'aménagements.
~
Les données de base furent recueillies du 30 août 1978 au
30 septembre 1979, dan~ fa zone démographique de l'Etang de
Miragôane, en Haïti, à environ 75 kilomètres à l'ouest de
Port-au-princ~par
20°39' de latitude nord et par 73°06' de longitudeouest. / ". 2. ,
,
,
,
l , i 1~(.
(
(~
Chapitre 1
Les caractères ~logiqUeS et la compétition des deux cichlidés de l'Etang de Miragôane,
~aiti,
CichZasoma haitiensis
Tee VanSa~~therodon
mossambicus
Peters.1
\
)
Soumis pour publication à la revue Hydrobiolosia
3.
.'
r
i
o
)
'..
- Cette première s'ection compare les deux cichlidés de l'Etahg
)
de Miragôane afin d'établir leur i~portance relative en fonction de la faune ichthyologique: Nous y comparons des paramètres de longueur, de masse, d'embonpoint ,~t de densité. Nous discutons également de l'introduction du'cichlidé africain en terme d'apport et de c~pétition à la population locale.D / / /
4.
\
5.
1.1 INTRODUCTION
1.1.1 Les antécédents.
Originaire de la côte est africaine,
S. mossambicus,
est introduit en Haïti en 1951, où 4 ans plus tard, il rep~ésente 90% des captures dans les eaux libres de la vallée de l'Artibonite (FAO lQ55). Pourl'Etang
de
Miragôane, il n'existe aucune étude scientifique sufrle succès de son implantation et de sa relation biologique avec lecichlidé
Cichlasoma haitiensis.
Chez ce dernier, i l n 'y a aucune référence quant à sa biologie.Par ailleurs, l'introduc~ion de
S. mossambicus
en Amérique latine a soulevé certaines polémiques liées à sa biologie et ·à la compétitionengendrée avec les c ichlidés indigènes (Myers 1955, Riedel 1965).
En fait Myers, suite à une revue de la faune ichthyologique d'Amérique
centrale, conclut que l'introduction de
S.
~os8ambicus aurait un effet, néfaste sur la productivité de la faune indigène. De plus il déclareque cette espèce apporte un bénéfice minime ou moindre à celui d'une
espèce endémique au point de vue de la produc,tivité. De son côté, '"
Riedel termine son étude des-cichlidés du Nicaragua en affirmant
1 '-apport bénéfique de l'introduction de
S. m08sambicus.
Il atteste qu'il ne saurait y avoir de compétition entre les adultes des espèceset qu'aucun cichlidé local ne peut rivaliser avec la croissance
rapide de
S. mossambiqus.
1.1.2 Les objectifs.
Suite aux contradictions entre Myers et Riedel, et suite à
dans un premier temps les caractères biologiques du cichlidé haïtien
:~
,1
et de S. mossambicus, afin cl 1 évaluer, le succès de l,! impla.ntation de
(
ce dernier. Pour ce, nous étudions les longueurs, 14] masses et les taux d'embonpoint spécifiques aux cichlidés de l'Etang de Miragôane.
'En second lieu, nous discutons la compétition inter-spécifique entre le ci~hlidé ha~tien et le cichlidé africain.
1.1.3 Le site (figure 4).
L'Etang de Miragôane se situe dans le département du Sud en
"
... .... ...--:::Haiti, a environ 75 kilometr~s à l'ouest de Port-au-Prince, par 20°39' de latitude nord, 73°06' de longitude ouest. Cette zone~xpérimen~ale
2
de 19.6 km , est un complexe lacustre formé de 3 plans d'eau reliés par une plaine marécageuse.
L 2 MATERIEL ET METHODES 1.2.1 Le matériel.
Essentiellement, le matériel d'échantillonnage consiste en des
~
filets maillants de type expérimental, composé de six panneaux de __ 7.6 mètres de lon-gueur chacun, associés à des mailles--é~irées de 2.5 cm, 3.8 cm, 5.1 cm, 6.4 cm, 7.6 cm et de lü.2,centimètres
(Ricker 1975).
1.2.2 Les méthodes.
1.2.2.1 L'enregistrement des données.
P,our
-!-'
ensemble des stations dans l'Etang de Miragôane, nous installons les filets pour une période de 24 heurea. Pour chaque poisson, nous enregistrons: l'espèce, -le sexe, la matu;ité sexuelle.6.
/
le contenu stomacal, le taux de parasiti;me, la longueur totale en
, 1
millimètres et la masse en grammes. Chez les individus sexuellement matures, nous pesons les gonades.
Par ailleurs, dans tout travail scientifique, l'auteur se voit confronté aux
ChOiX~S
tesfs statistiques adéquats. Il se doit d'utiliser "l'ou~proprié
à ses besoins et à ses contraintes.Il
Dans notre cas, nous délaissons les tests statistiques paramétriques lorsque nous constatons la violation prononcée des assomptions de base. Alors nous utilisons les modes et les médianes comme mesures de tendances, centrales. Pour l'ensemble des tests statis tiques,
nous adaptons un seuil critique égal à 57..
1.2.2.2 L'étude de la longueur.
Pour l'ensemble des longueurs, nous établissons les histo-grammes de fréquences selon l'espèce et le sexe. Nous divisons ces histogrammes en groupes où nous y regardons la distribution des individus selon leur densité et leur sexe. Ces groupes relèvent des modes issus des histogrammes de fréquences de longueur. Ces derniers sont considérés en ichthyologie comme des indicateurs des groupes d'âge de la population, où certains peuvent toutefois ssimiler plusieurs âges (Ricker 1975). Cette analyse identifie
longueurs représentatives des populations de cichlidés, éprouvées
par des tests de médianes, de Kruskal-Wallis et de Chi-carré (Siegel 1956).
1.2.2.3 L'étude de la masse en fonction de la longueur.
Nous considérons les informations issues de la masse spécifique des 2 cichlidés via les relations de Longueur-Masse, sous la forme
de régressions logarithmiques du modèle suivant (Ricker 1975): Log M = Log a + n Log L
/ où M représente la masse en grammes, a l!ordonnée à l'origine,
---/
n la pente de la régression et L la longueur totale du poisson. Nous comparons ces régressions entre les populations d'Haïti ainsi - qu'avec celles des populations de S. mossambiaus issues de régions tropicales et néo-tropicales telles que le Nicaragua, l'Egypte et Hong Kong.
La différenciation statistique de ces régress,ions s'effectue par des tests de t sur les pentes. Quoique la violation des assomptions de base affaiblit ces tests, les conclusions hautement /, significatives (p <0.01) demeurent valides vue la puissance même
.
c!.Y/
test et vue la valeur élevée du coefficient de corrélation ,attaché--généralement aux relations Longueur-Masse
1.2.2.4 L'étude de l'embonpoint.
Le coefficient de condition ou d'embonpoint, K, permet d'évaluer la "santé" d'une population par le rapport entre la masse et la longueur de l'individu selon la formule (Ricker 1975):
où M représente la masse totale du poisson en grammes et L la longueur totale en millimètres. La forme commune des cichlidés permet de comparer le coefficient de condition des 2 espèces afin de distinguer le degré de succès dans l'assimilation des ressources pour S. mossambiaus. Aussi nous réunissons les modes de longueur grâce aux valeurs de K, éprouvées par le test de Kruskal-Wallis,
/
../
pour former des classes d'individus telles que les juvéniles, les immatures et les adultes (figure 3).
:.' :~: ' ~, .;j ~\
1
1. 3 RESULTATS
Les paramètres biologiques des cichlidés de l'Etang de Mifagoâne.
1.3.1 Sarotherodon mossambiaus.
1.3.1.1 La longueur de S. mossambicus (figure 1).
1
L'histogramme des fréquences de longueur 111ustre 6 modes dans la population. Suite" à ceux-ci, nous délimitons 6 groupes de
longueur: 60-95 mm, 96-125 mm, 126-155 mm, 156-185
mm,
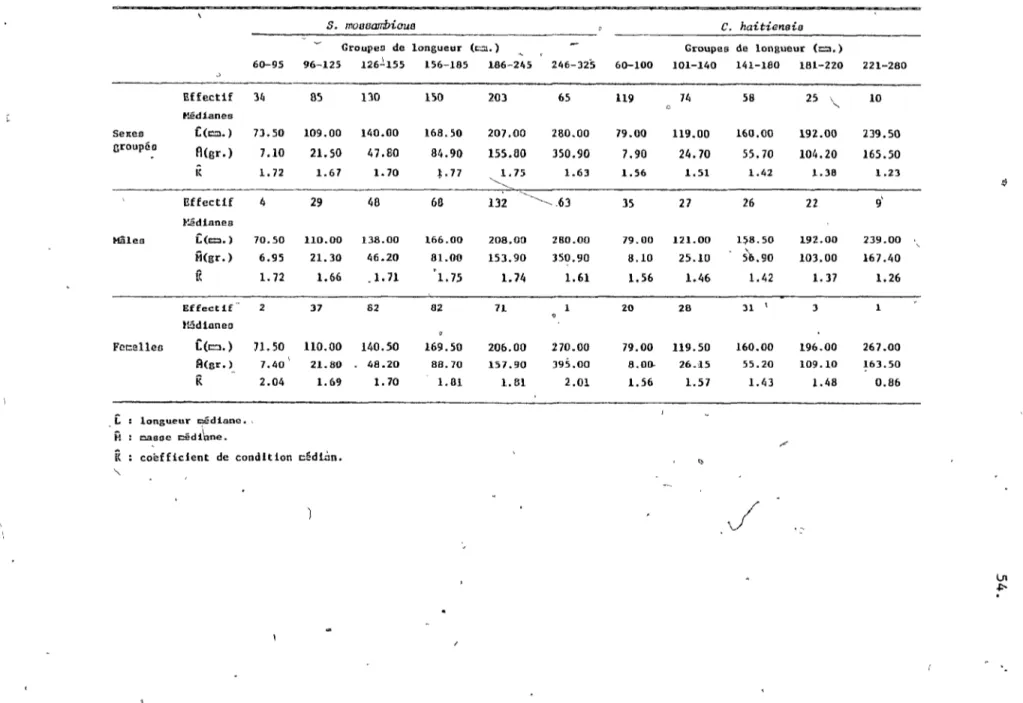
186-245 mm, 245-325 mm. Globalement, il y appert que plus de 70% des individus mesurent entre 126 mm et 245 mm tandis que seuLement 5% indiquent une taille inférieure à 95 millimètres.La composition sexuelle de la population, soit le nombre de
1
mâles et de femelles, varie en fonction de la longueur CP <0.001).
?
Ainsi nous observons les rapports mâles
1
femelles suivants: 96-125 mm, 1:1 (p >0.05); 126-185 mm, 5:7 (p <0.001); 186-325 mm, 3:1 CP <0.001). Alors dans son ensemble, la population de S. mossambiaus démontre bien la dominance en taille et en nombre des mâles (tableau 2).1.3.1.2 La masse de S. m08sambicu8 (figure 2),
Nous établissons pour cette population, une relation de Longueur~
Masse définie par l'équation suivante: -5
Log M
=
Log (1.98 X 10 )+
2.9720 Log L" ,
10.
Selon les sexes, nous obtenons des régressions différentes pour chacun:
Mâles: Log M
=
Log (2.25 X 10-5)+
2.9468 Log L,
Femelles: Log M = Log (1.55 X 10-5) + 3.0233 Log LEn fait, la croissance en masse des femelles s'avère supérieure à
J 0
celle des mâles, tel qu'indiqué par les pentes (p <0.001). Néanmoins, les mâles possèdent une masse médiane dominante
CP
<0.001).1.3.1.3' L'embonpoint de S. , mossambicus (figure 3).
L'illustration graphique du coefficient de condition en fonction de la longueur, démontre des variations parmi les groupes de longueur.
, b
Ainsi les longueurs 60-95 mm et 155-245 wm pospèdent un K médian
commun (p >0.05). Les autres, 96-155 mm et 246-325 mm se qualifient d'un· K médian semblable (p >0.05) mais inférieur aux autres groupes (p <0.05). Alors selon ces caractères biologiques nOtl~~éfinissons 4 classes de longueur dans la population de S. mossambiaus;
60-95 mm les juvéniles 96-155 mm: les immatures 156-245 mm: les adultes mixtes 245 mm et plus: les mâles aduLtes
1.3.2 Cichlasoma haitiensis.
1
1.3.2.1 La longueur de C.
haitiensis
(figure 2),Chez ce cich1idé indigène, l'échantillonnage de la population révèle 5 modes, d'où la délimitation de 5 groupes, soient 60-100 mm, 101-140 mm, 141-180 mm, 181-220 mm, 221-280 mm, où plus de 85% des
"
11.
poissons mesurent moins de 180 mm. Parmi ces groupes, la répartition des mâles et des femelles varie
Cp
<0.001). Néanmoins, ceux de60-100 mm, 101-140 mm et 141-180 mm, in,\~quent des propol;tions
"-communes
Cp
>0.05),soit 1:1 (p >0.05). Globalement les mâlesdominent donc la population de par leur nombre et leur taille (p <0.05) (tableau 2).
1.3.2..2 La masse de C. haitiensia (figure 2., tableau ~).
'La relation
"LOngUeUr-Mass~
cette population s'illustre selon cette formule:Log M =: Log (3.56 X 10-5)
+
3.8136 Log LDt autre part:', la cr9issance en masse s'avère supérieure è.hez les
mâles
CP
<0.001). suivant les régressions:Mâles: Log
M~==
Log (3.62~
10-5)+
2.8109 Log L Femelles: Log M = Log (5.34 X 10-5)+
2.7327 Log LAussi, la masse médiane des mâles est plus grande que celle des~ femelles Cp <0.001).
1.3.2.3 L'embonpoint de C. haitiensis (figure 3).
Chez cette population, nous remarquons une diminution constante du coefficient de condition lors de la croissance en longueur.
L'analyse du K selon les groupes de longueur, réunit ces derniers en classes. Ainsi celle de 60-140 mm possède un K médian commun
.---:-Cp >0.05) et supérieur auX autres classes (p <0.05). La classe
intermédiaire assimile les groupes 141-180 mm et 181-220 mm. La dernière, 221-280'mm, dénote une valeur minimale de K pour des
mâles mat ures,
/ '
\
\
\\
\
\ \1.3. 3 Paralle~es biologiques entre ces cichlidés. 1.3.3.1 La densité (tableau 1).
12.
(
Pour l'ensemble, S. mossambiaus affiche une supériorité numérique de l'ordre de 2 pour '1 C.
haitiensis
(p <0.001). Par contre, la proportion de ~lest.t
de femelles ne change pas selon les espèces"
(p >0.05), où notls rèma~quons de 25% à 40% plus de mâles que de
f\
femelles.
1.3.3.2 La longueur (figure l, tableau 1).
Globalement, les histogrammes de fréquences et la comparaison des longueurs, démontrent une taille supérieure, chez ,mo?sambiaus, de 46% à celle de
C. haitiensis
(p <0.001). Ceci seles modes principaux, où pour S~ mossambicu8 les deux odes princ~paux sont 160-170 mm et 200-210 mm, et où pour C.
haitiensis
le modemaj eur es t 70-80 ~.
taille
l
sUPérie\Ure à
,
Chez les deux espèces, les mâles possèdent une
...
celle des femelles.1.3.3.3 La masse (figure 2).
Dans l'Etang de Miragôaoe, les deux espèces possèdent leur
propre croissance en masse, où
S.
mossambicus y affirme sa suprématie"
"
Cp <0.001). Par contre, le taux de croissance en masse de S.
mossambicus est inférieur à ceux des populations étrangères.
Cependant, vu l'absence des paramètres statistiques liés aux régressions, ces comparaisons demeurent sujettes à caution.
La masse spécifique de
S.
mossambious est de 4 fois supérieure à celle deC. haitiensis
(p <0.001) (tableau 1)...
. /
.
(
, ,/
1.3.3.4 L'embonpoint (tableau l')
La m~sse spécifique de S. m08sambieus engendre un coefficient diembonpoint de 17% supérieur à celui du cich1idé haïtien.
1.3.3.5 Des observations dive"rses
Des observations régulières sur le terrain apportent des
.)
précisions SU4!" 'la biologie des 2 espèces. Danb le cas de la nutrition,
nous notons l'utilisation de 2 ressources principales. A~nsi chez
S. mossambieus, les individus forment des bandes et ils filtrent le
phytoplancton dans les eaux de surface des zones d'eau libre. Chez
C. haiJien,sis la diète alimentaire se base principalement sur les
_ /
algues ChQ1'as sp., tel qu'observé -dans l'estomac des individus
capturés. Ces individus possèdent aussi un taux élevé de parasites internes et externes". Cette espèce affiche en p,lus des caractères néoténiques, relevés lors de la dissection des captures, p~incipalement la maturation des gonades.
1.4 DISCUSSION
1.4.1 Le succès de S. mossambicus.
Actuellement, les Rrises de S. m08sambicus représente plus de 70% des captures de 1 'Etang~e Miragôane. Il forme une pqpulation ichthyologique stable et en bonne condition, tel qu'indiqué par , la valeur spécifique de K. Cette espèce se répartit en 4 catégories
D 1 \
d~~.~'dividus:
les juvéniles, les immatures. les adultes mixtes, les mâles adultes. Les juvéniles correspondent aux indivtdus inférieurs
t-(
14.
caractéristique retrouvée chez les cichlidél:l (?Riedel 1965, Bowen
1979c). Elle provient de l'utilisation des terrasses et de Il eulittora1 . comme pouponnière (Lowe 1952, 1958, Fryer et Iles 1972, Bruton et
BoIt 1974, Whitfield et Blader 1978a,b) (cf. chapitre 2).
De 96 mm à 155 mm,.. nous retrouvons des poissons majoritairement immatures, qualifiés d'un K minimal. Cette baisse du coeffiçient de condition provient du st~de d'immaturité sexuel et aussi de ,l'utilisation d'un milieu différent, les
- ~
-'Allanson 1974, Bruton et Bo1t'-,1974) (cf.
zones d' eat libre (Brut on et
~
chapitre 2)"puisqu~
cetteD
zone s'avère être de qualité nutritive inférieure causée par le manque de diatomées (Lowe 1958, Bowen 1976, 1978,1979 a,b,c).
1
Les adultes mixtes. d~ 156 . mm à 245 mm de longueur, possèdent des coefficients de condition élevés et statistiquement équivalents à ceux des juvéniles. Cette condition dérive de la maturation des gonades. couplée au retour dans les terrasses (Lowe 1958. Bruton et BoH 1974) (c.!, chapitre 2).,
Les mâles adultes, de longueur supérieure
-
à 245 mm se distinguent
-par un coefficient de condition faible. Cet état relève du sommet de croissance atteint par ces individus et de l'occupation dl une
i
zone de qualité nutritive faible (Riedel 1965)
Ccf.
chapitre 2). Cette population suit donc le cycle biologique propre à son espèce, tel que décrit dans les autres régions tropicales et néo-tropicales. quoiqüe sa croissance en masse y soit légèrement plus faible.i
..
/',
1 )
)
(
,-
--,1.4.2 L'infériorité de C. haitiGntsis et la compétition avec
S. mossarribicus.
La population du cich1idé haïtien accuse une infériorité marq uée sur celle du cich1idé introduit, tant' du point de vue de densité que de longueur, de masse ou d'embonpoint. Cet état découlerait-il de la compétition avec S. m08s(JJTlbicus?
Tout au long de l'étude, le contenu stomacal des adultes et des immatures demeure constant et démontre l'utilisation quasi-exclusAve des characées comme aliment pour C.
,
haitiensis. ChezS. mO~8ambicus, les adultes et les juvéniles se nourrissent
principalement de phytoplancton et d' aggrégats périphériques de détritus retrouvés sur les macrophytes (Bruton et BoIt 1974, Bowen 1979, 1979b,c, LeRoux 1956, Lowe 1958, Whitfield et Blader 1978a,b). Alors à ces stades. ces espèces ne rivaIisent pas pour les sources nutritives.
Par contre, les alevins et les juvéniles peuvent concurr~ncer. A ce stade ils utilisent vraisemblablement des sources nutritives communes, exploitées par
i'
ensemble des cichlidés, soient les dépots flocculeux répandus sur le fond de l' eulittoral et très riche endj.atomées (Bowen 1976, 1979b,c, Bruton et BoIt. 1974). Toutefois cette ~ériode de rivalité se trouve restreinte principalement aux équinoxes de printemps et d'automne où les pluies stimulent la fraie de
S. mossambicus (Hodgkiss et Man 1978, Riedel 1965).
Dès lors, la compétition alimentaire ne suffit pas à expliquer
"
l'état médiocre de la population de C. haitiensis. En fait plusieurs indices penc~ent plutôt vers l'hypothèse de la surpopulation. En
1 ...
(
,eff et, la res sou,rce alimentaire très spécifique et t rès lo~ale que sont les characées, le taux élevé de parasitisme et la néoténie observée sur le terrain sont autant d'indicateurs d'une population restreinte par le milieu utilisable (Fryer et Iles 1972).
1.5 CONCLUSIONS
L'ensemble de cette étude corrobore les affirmations de Riedel, démentant par le fait même celles de Myers. L'introduction de
-.,
S. mossambicus ne peut causer de préjudice sérieux -à la population
\
, indigène, d' aille~rs de faible productivité, puisque cette dernière,
à l'état adulte, se nourrit de characées et que S. mossambicus est
---~
principalement phytoplanctophage à ce même stade.
La croissance rapide de S. mossambicus se traduit dans la
16.
population des cichlidés par une densité accrue, une taille supérieure, une croissance de masse plus rapide et un coefficient' cl' embonpoint, plus élevé. Alors l'introduction de ce cichlidé entraîne une
augmentation tangible de la productivité de la faune ichthyologique
---de l'Etang de Miragôane, où i l devient l'espèce dominante. Cette dominance de biomasse par S. mOBsambicu8 provient de l'utilisation d' une ressource majeure et inutilisée par son homologue ha1tien, soit :j.e phytoplancton des eaux libres.
Pour terminer, le t4ux élevé de parasitisme chez C. haitiensis
ainsi que la néoténie observée chez les 2 espèces, pourraient indiquer une ressource ichthyologique sous-exploitée.
, .,
)
17. ' ..'.,
'.
-, ;.
Ch-:pitre 2Analyse descriptive de l'Etang de Miragôane,
Haïti, et répa;tition des cichlidés,
CichZasoma haitiensis
Tee Van etSarotherodon
mO~8ambicus Pet~rs,en fonction des milieux.
Soumis pour publication à la revue Hydrobiologia
18.
Suite à l'évaluation du potentiel des deux cichlidés, il importe
~
de déterminer l'importance relative des habitats retrouvés à
l'intérieur de l'écosystème marécageux de Miragôane. Il s'agit
d'analyser la répartition spécifique des cichlides en fonction des
lacs et de la végétation. Ces informations, reliées à la biologie
des espèces, permettront de définir les zones lacustres essentielles
au maintien .de la productîvité ichthyologique de l'Etang de
Miragôane.
j 1
2.1 INTRODUCT ION
2.1.1 Les antécédents.
-
"-En 1951, la Food and Agricultur~ Organization
(FAD)
introduisa~t un cichlidé africain, Sarotherodon mossambicu8, dans les eaux libres d'Haïti (Lin 1952). Depuis, ce cichlidé a démontré le succès de son implantation. Il, surpasse la densité el la croissance en masse de son homologue haïtien, Cichlasoma haitiensis, sans engendrer de~ 1
compétition inter-spécifique
(ci.
chapitre 1). Le succès d'adaptation de ce cich1idé dépend en grande partie d'une ressource inexploitée par le cichlidé haïtien: le phytoplancton.De nos jours, il est démontré que les migrations inter-habitats dans les eaUX lacustres sont à la base de l'écologie des cichlidés. En fait, ces poissons utilisent des zones de faible profondeur pour la reproduction et l'élevage des alevins (Lowe 1955, Welcomme 1964, Bruton et Allanson 1974, Bruton et BoIt 1974, BOHen 1979a,b,c). Puis les poissons se dispersent dans les milieux pour profiter des ressources propres aux différents lacs. De par ce point commun aux diverses
populations de cichlides, Welcomme (1964) qualifia le nombre de ces zones d'élevage comme le facteur limitant la productivité ichthyologique tropicale.
2.1.2 Les objectifs.
Suite aux COll i SSances trop générales sur l'écologie des
..
c(iChlidés, nous analysons ici l'utilisation des zones lacustres del'Etang de Miragôane afin d'évaluer l'importance relative de ces milieux aquatiques en fonction du cycle biologique des cichlidés .
Ces nouveaux renseignements~pourraient, permettre un aménagement adéquat de la région.
L'étude de l'activité inter-habitat concerne les deux espèces
/
20.
de cichlidés réparties dans les plans d'eau et les zonftS du littoral, où nous y comparons des paramètres biologiques tels que la longueur, la masse et le coefficient de condition afin d'identifier les groupes ichthyologiques occupant ces milieux.
2.1.3 Le site (figure 4).
L'Etang de Miragôane, est un des principaux plans d'eau douce en Haïti.
Il
est localisé à l'ouest de Port-au-Prince par 20°39' de latitude nord et 73°06' de longitude ouest.- Il occupe une superficie
~-2
de 19.6 km et il englobe différentes associations pedo-végétales,
1
}) agricoles Ou naturelles.
2.2 MATERIEL ET METHODES--~'
2.2.1 Geographie physique.
Nous cartogr'aphions la 17égion à l'aide de photos aériennes de la serie 78-HA1-Ol/400 UAG 412, de numero 1009 - 1010 - 1011 - 1012 et d~éche1le 1:50,000 ainsi qu'avec les cartes topographiques de
Petit Gôave et de Miragôane d'échelle 1:50,000 et de numero 5671-11-111. Ces outils proviennent du Ministère Haïtien dT Géographie et de
Cartographie. Pui$ nous identifions ces macrozones de vegetation lors des visites sur le terr~in. Nous analysons la végetation aquatique,à l'aide 4e,transects perpendiculaires à la rive, sur
des distances régies par la présence des macrophytes, lors de plongées en apnée. Nous utilisons un gradient empirique de 1 à 4, pour
quantifier la densl:.:é relative de la végétation.
Selon les cartes morphométriques des plans d'eau, effectuées par la firme d'ingénieur conseil Sohic~o, nous parachevons l'étude physique des lacs pour évaluer leur volume et leur profondeur moyenne
(Wetze1 1975). Notons que les surfaces étudiées au cours de ce travail sont évaluée selon la technique du "Dot 'Gr id" .
.-/
2.2.2 Analyse physico-chimique de l'eau (Wetzel 1975).
A l'aide de l'échantillonneur à renversement Nanson, nous déterminons les profils de ~ifférents paramètres physico-chimiques. Pour le taux d'oxygène dissous nous adoptons la métliode de titration de Winkler. Le laboratoire du Département de l'Agriculture des Ressources Naturelles d'Ha'tti effectua les analyses du pH, de la
\
cônductivité électrique, des cations, des nitrates et des ions
carbonates. L'emplol du disque de Secchi caractérise la transparence de l'eau, dont le double de la mesure indique la profondeur limite-de la zone euphotique.
2.2.3 _Echantillonnage ichthyologique.
Des filets mail1ants de type expérimental (cf. chapitre 1), sont installés parallèlement dans les divers habitats aquatiques pour des périodes d'effort égales à 24 heures. Nous adoptons les mesures de longueur totale en millimètres et celles de masse en grammes. Puis nous évaluons le coefficient de conditions. K des poissons (cf. chapitre 1).
!
22.
Pour l'ensemble, nous fixons un seuil statistique critique de 5%.
Les tests statistiques paramétriques sont délaissés au profit des épreuves non-paramétriques car les données ne rencontrent pas les assomptions de base~requises par les outils statistiques paramétriques. Ainsi le test de Chi-carré vérifie les analyses de fréquences et les épreuves de Kruskal-Wallis comparent les mesures de tendances centrales (Siegel 1956).
2.3 RESULTATS
2.3.1 L'Etang de Miragôane (figure 4). 2.3.1.1 Caractêres physiques.
Le marais de Miragôane comporte 3 lacs associés à un bassin de
2
drainage de 149 km Le Petit Etang y occupe une superficie de 2
1.2 km avec une profondeur maximale de 22 mètres, une profondeur moyenne de 11 mètres et un volume de 1. 3 X 104 m3. L'Etang de Miragôane se divise en 2 sections, A et B. La partie sud, le
2
lac A , d'une surf ace de 8.2 km , acc use une profondeur maximale 8 3
de 43 m, un volume de Z. 3 X 10 m et une profondeur moyenne de 28.4 m. Selon l'analyse physico-chimique de l'eau (figure 5), le lac A se qualifie cl 'oligomictique à profil d'oxygène clinograde, où à une profondeur supérieure à 28 m, nous y remarquons la présence
olfactive du sulfure d'hydrogène, HZS. libéré lors de la décomposition
: / ,"
anaérobique des sédiments organiques (Wetzel 1975).
La section nord de l'Etang de Miragôane se définit par une superficie de 0.9 km2, associée à un volume de 793 m • et une 3
./
/
---~--~
..
23.
2.3.1.2 Les associations végétales.
La section marécageuse de l'Etang de Miragôa~e se caractérise par différentes colonisations végétales. La majorité des secteurs supporte une culture mixte de tubercules et de céréales d'une étendue
2
globale de 5.3 km Elle repose sur des sols argileux hydromorphes, sauf pour le versant sud, caractérisé par des sols limona-argileux à pseuaogley de drainage imparfait (D.R.I.P.P. et Thai 1978).
La seconde zone d'importance, les marais de cladiums, occüpent une superficie de 2.8 km2. Ils possèdent des sols tourbeux mal décomposés de drainage nul. En dernier lieu viennent les marais
d'éléo~harides, de surface égale à 1.6 km2 • Ceux-ci occupent en
fait le littoral supérieur de l'Etang de Miragôane.
La zone euphotique du lac, soit le littoral de niveau d'eau inférieur à Il mètrés, s '~appropriè 14% de la superficie lacustre,
[ ,
soit 1.1 km2 (figure 6). Selon la profondeur, il y apparait certaines associations végétales. Ainsi les marais de cladiums,
CZaaium
jamaicensis,
y colonisent l'eulittoral. - Puis le littoral supérieur,~e des éléocharides,
EZeooharis interstinata, EZeocharis
robinsii~évolue dans le littoral moyen en une association de rubaniers,
VaZisneria
amerioana~ et de 'potamots,Potamogeton iZionensis:
Celle-ci2
plus de la moitié du littoral actuel, soit 0.6 km . accapare
Finalement, le littoriprofondal révèle une pente croissance et il supporte quelques potamots mais majoritairement des Naias de la Guadeloupe J
Naj
as guada ~upen8i8.
1
"
une couverture végétale pour pl~s de 75% du fond. Les associations
s'y divisent en 3 groupes homogènes, les potamots, les rubaniers
et les characies.
1 2.3.1. 3 Les milieux.
Suite à des caractéristiques physiques et écologiques, nous
réunissons différentes stations susceptibles d'influencer la
distri-bution des cichlidés. Trois milieux sont ainsi définis, en fonction
de la profondeur et de la végétation aquatique (figure 6). Le
milieu l correspond aux terrasses 'du littoral où la végitation couvre
'.1
à 100% le fond du lac, par une profondeur de 1 m à 2 m. Le couvert
/
24.
végétal se compose majoritairement de potamots,
Potamogeton ilionensis
Morong, et de rubaniers, Valisneria americana Michx.
Le milieu II regroupe une section du littoral moyen et du
littoral inférieur où la couverture végétale garnit à 75% le fong
du pla~ d'eau. La profondeur de ce milieu varie de 2 mètres à 4
mètres. La composition végétale comporte pour le littoral moyen,
une association dominante de potamots avec une présence régressive
.---de rubaniers. Dans le littoral inférieur, les characées et les
naïadacées,
Najas guadalupensis
Spring,dominent.f •
_ / >
Le milieu III coalise toutes les stations où la couverture
végitale'- est inférieure à 25% et se compose principalement des naïada,cées
,
j
associées aux characées. La profondeur dépasse 4 mètres. Nous
classifions cet habitat de zone d'eau libre.
~ 25. /
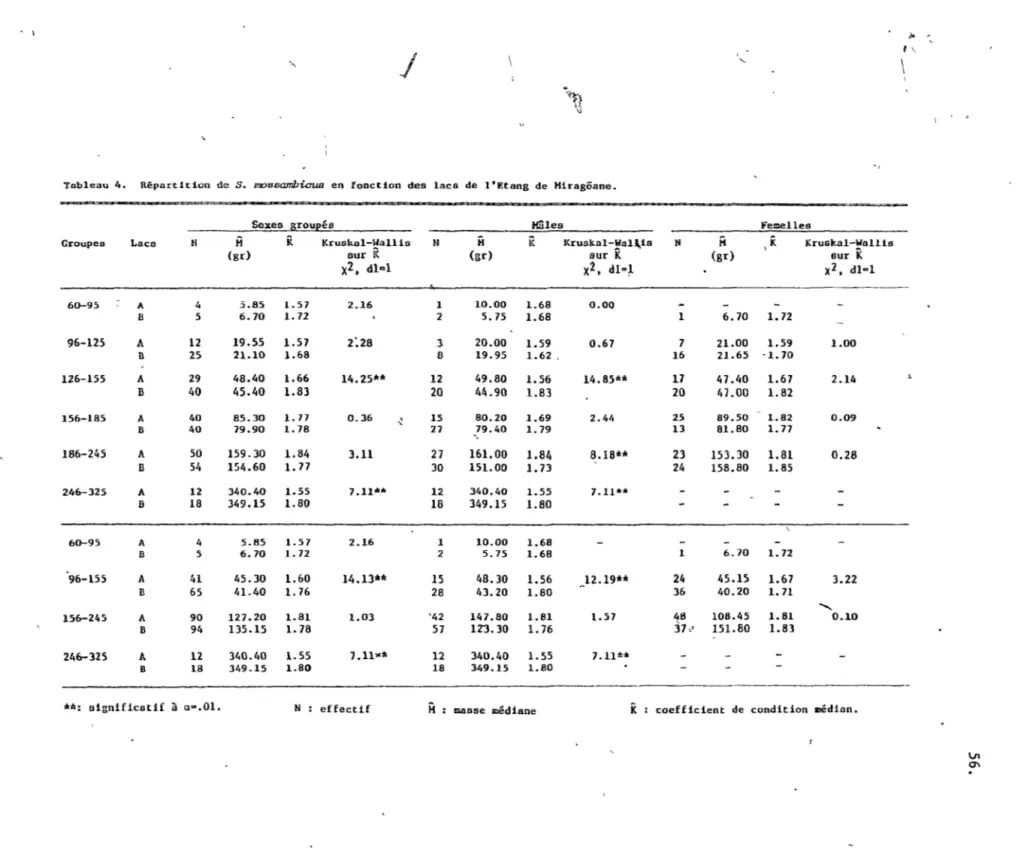
2.3.2 La répartition des cichlidés en foncti'on des plans d'eau.
2.3.2.1 SarotheT'odon m088ambious (tableaux 3 et 4).
Pour les fins de cette étude, nous divisons cette population en six groupes de longueur, soient 60-95 mm, 96-125 mm, 126-155 mm,
156-185 mm, 186-245 mm, 245-325 UIDl, et en 4 classes soient les
juvéniles, les immatures, les adultes mixtes et les mâles, adultes
(cf. chapitre 1).
Chez cette espèce, sans égard au sexe, nous observons des populations égales en taille entre les deux lacs, que ce soit selon les groupes de longueur ou selon les médianes de longueur (p >0.05). Not'ons t~ut(.fois un coefficient de condition supérieur
pour celle du lac B
Chez le's mâles, la même analyse révèle une concentration de ceux-ci dans le lac B selon le rapport de 3:2, (p <0.01) quoique les longueurs et les masses médianes des deux populations ne . diffèrent pas. Cependant celle du lac B comporte des mâles plus gras, spécialement parmi les groupes 126-155 mm et 246-325 mm.
Chez les femelles, nous observons dans l'ensemble une distribution homogène des classes' de longueur en fonction des lacs (p >0.05).
Néanmoins, les p.oissons des groupes de longueur 60-95 ~ et 96-155 mm
se concentrent dans le lac B selon un rapport 7: 5 (p < O. 05). Par contre, aucune différence significative ne se rattache aux populations de femelles des 2 lacs, quant aux médianes de longueur, de masse et d'embonpoint.
t
~2.3.2.2 Ciahlaaoma haitiensis (tableaux 3 et 5).
Selon 1 'histogramme des fréquences nous délimitons 5 groupes
l
de longuel'r; soient 60-100 mm, 101-140 mm, 141-180 mm, 181-220 mm, 221-280 mm, qui sont regIioupés par la suite selon leur coefficient de condition (cf, chapitre 1).
Globalement, C. haitiensis se retrouvé irrégulièrement parmi
~
les lacs en fonction de la longueur (p <O. al). En fait les petits individus, de 60 mm à 140 mm de longueur, se concentrent dans le lac B selon des proportions de 6:1 Cp <0.001), Les autres, de taille supérieure à 141 mm, se distribuent uniformément dans les eaux des lacs A et B (p > 0.05) . Par ailleurs, cette préférence des petits ind}vidus pour le lac B se réflète dans les paramètres médians de lo~gueur et de masse, où celui de
K,
supérieure pour le26.
lac B , révèle l'influence bénéfique de ce choix POlfT les juvéniles, 60-140 mm.
La même analyse répétée en fonction des sexes indique toujours
/
.une préférence des petits individus pour le lac B (p <0.001).
\
Ceci se concrétise par l'assignation très signif~cative de médianes supérieures, de longueur et de masse, pour la populat ion du lac A .
,
...
De plus, notons chez les femelles, une valeur de K nettement supérieure pour le lac B, attribuable vraisemblablement à une
\
..
(.
~
\ \ 2.3.3 La répartitionde~~'enJonction
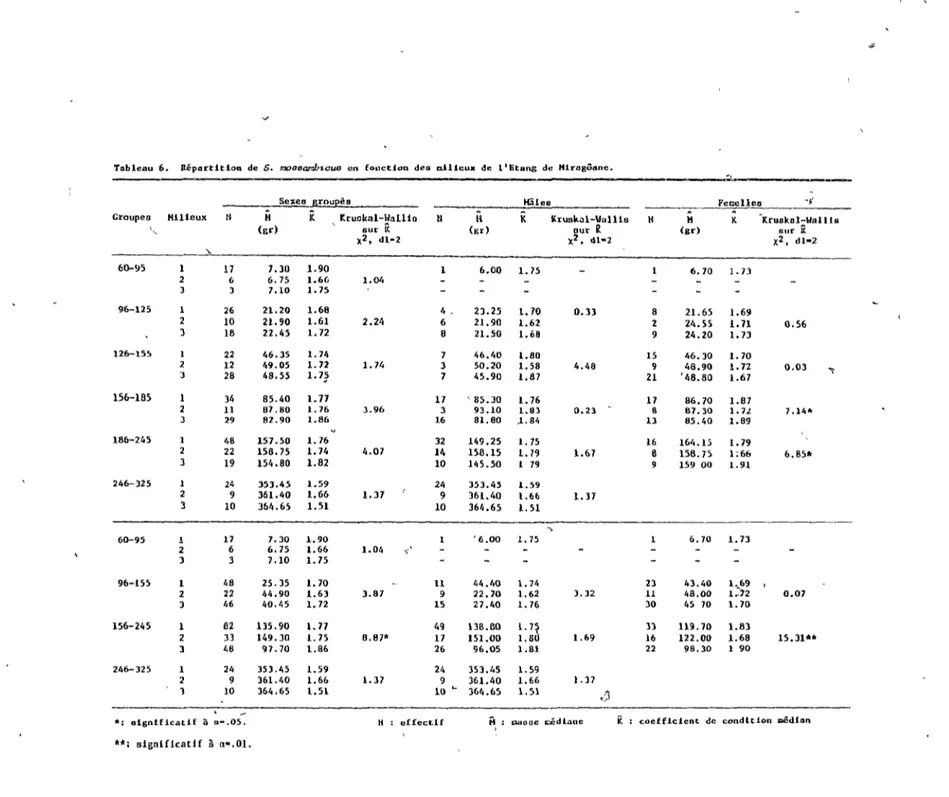
'\2.3.3.1 S. mossambicus (tableaux 3 et\6).
des mi"1ieux.
Dans son ensemble, la répartition det groupes de longueur en fonction des milieux se révèle non uniform~(p <0.01). Les poissons
\ 1
de taille -I.nférieur à 95 mm, indiquent une priÜérence pour les milieux de végétation dense selon les proportions 6:3:1 (p <0.01). Ceux de longueur variant entre 96 mm et 155 mm, se retrouvent uniformément dans les 3 milieux Cp >0.05). Chez les individus
-supérieufs à 156 mm, nous notons une sélection des milieux à
végé tation dense dans l'ordre de 12: 2: l (p <0.001).
Chez les mâles, ceux d~ taille inférieure à 185 mm semblent se distribuer de façon homogène parmi les milieux (p >0.05). autres, de 186 mm à 325 mm de longueur, se concentrent dans les
"
milieux de végétati~n dense selon le rapport 3:2:1 (p <0.001). D'ailleurs, la population du milieu III possède des médianes de longueur et de masse nettement inférieures aux autres milieux .
.
Chez les femelles, nous remarquons une distribution_homogène en fonction des milieux (p > O. 05) . Notons cependant un K supérieur ,pour les femelles du milieu III, attribuable à la classe 156-245 mm.
Ainsi, l'analyse de la distribution des individus parmi les milieux, en fonction des sexes, ne révèle pas de préférence des petits indiv~dus pour des milieux-de végétation dense. Ceci dépend
1
de l'impossibilité de déterminer convenablement le sexe des poissons mesurant moins de 95 millimètres, nous obligeant à ignorer le
g~oupe 60-95 mm pour cette analyse.
27.
28.
2.3.3.2 C. haitiensis (tableaux 3""et 7),.
Ce cichlidé dénote uné répartition dissemblable des groupes de longueur (p <0.05). Le groupe ,60-100 mm se concentre dans les eaux libres selon le rapport 2:!:1 (p <0.001). Les autres groupes observent une distr~bution homogène parmi les milieux (p >0.05).
Chez les mâles, nous observons pour l'ensemble des classes deux stratégies, un premier groupe dans le milieu l et un second dans les eaux libres (p <0.01). Chez le~ femelles, les groupes 60-100 mm et
101-140 mm se distribuent parmi les milieux de la même façon que les mâles. C'est-à-dire qu'elles délaissent le milieu II. Les autres
/ '
groupes ne présentent aucune différence de densité parmi les milieux (p >0.05). Les médianes indiquent par ailleurs une population
-, inférieure en -longueur et en masse poui: le milieu III, mais dotée
d'un
K
supérieur.2.4 DISCUSSION
/
2.4.1 S.
mossambious.
Chez cette espèce, la répartition de la population parmi les habitats dépend de la taille des individus et du sexe. Globalement, les mâles colonisent surtout la section B de l'Etang de Miragôane, couverte à plus de 75% par les macrophytes. Cette collectivit~ y accuse aussi un coefficient d'embonpoint supérieur aux congénères du lac A. Parallèlement, l'analyse en fonction des milieux indique une préférence des mâles pour les terrass'es supérieures. Ces mâles
(
.1
(
29.
correspondent principalement à la classe 245-325 mm, composée de mâles matures défendant un te,rritoire de reproduction (cf. chapitre 1).
Chez les femelles, la distribution des individus s'effectue uniformément parmi les lacs A et B et·parmi les habitats analysés. Cependant celles occupant le milieu III, accusent un coefficient d'embonpoint supérieur à celui des autres milieux. Cette condition dérive -de l'utilisation des eaux libres, riches en phytoplancton et plus exposées au soleil, afin de compléter la maturation des gonades
\
(FAO 1955, Bruton et Allanson 1974, Bruton et BoIt 1974, Bowen 1976, Whitfield et Blader 1978a,b, 1979). Ainsi l'augmentation de poids attri-buable aux ovaires matures contribue à augmenter significativement la valeur du K pour cette classe d'adulte de tail.le variant entre 156 mm et 245 mm (cf. chapitre
1).
Chez les individus non-reproducteurs, 60-lf5 mm, nous observons une concentration dans le lac B. Par contre, l'analyse selon les habitats décèle deux tendances parmi ce groupe. Ainsi, les juvéniles,
of, 60-95 mm (cf. chapitre 1) démontrent une densité supérieure dans le
milieu l, soit la partie supérieure des terrasses, nantie du couvert végétal le plus dense et d'une profondeur inférieure à deux mètres. Chez les jeunes immatures, 96-155 mm (cf. 'chapitre 1) nous observons l'homogénéité parmi les milieux.
De cet ensemble de caractéristiques,' il appert donc une sélection des habitats lacustres en fonction du couvert végétal. En fait, les juvéniles, 60-95 mm de longueur (cf. chapitre 1) se concentrent dans le littoral supérieur où ils profitent d'un abri végétal adéquat et d'une diète plus complète (Welcomme 1964, Bruton et Bolt 1974, Bo'Ten 1976, 1979c).
1
)
(
,7
Au stade de jeunes immatures (cf. chapitre 1), ils se dispersent
-~ans l'ensemble de l'Etang de Miragôane pendant la maturation des
gonades. A maturité sexuelle (cf. chapitre 1) ils retournent dans
les terrasses pour y frayer et y élever les alevIns, qui seront
libérés par la suite dans l'eulittor~l (Lowe 1958, Bruton et A11anson
1974,
Bruton et BoIt1974).
Une fois l'élevage terminé, d'une~duréed'environ 2 semaines (Des texhe-G~mex
f967).,
les' de nouveau, tandis que les mâles demeurent dansfemelles se dispersent
le littoral_moyen pour
y défendre un territoire et pour s'y accoupler avec d'autres femelles.
2.4.2 C.
'haitiensis.
Chez celui-Ci, les deux sexes préfèrent, selon leur état de
maturité sexuelle, ou les ,terrasses, ou les zones d'eau libre,
sui~t le schéma décrit précédemment. Toutefois, l'importance sélectif du lac B de l'Etang de Miragôane prime sur le gradient
des milieux, pour ce cichlidé. L'explication à ce phénomène relève
de la forte densité ae characées dan~ le lac
B.
Ces alguesconstituent en fait la principale source nutritive de cette espèce
(cf. chapitre 1).
2.5 CONCLUSION
Les cichlidés de l'Etang de Miragôane effectuent des migrations
inter-habitats au cours de leurs cycles biologiques, de façon à
satisfaire aux besoins des divers stades. Ainsi ces poissons
31.
d'élevage soient principalement les marais de cladiums et d'éléocharides
ainsi que l'association de potamots-rubaniers. De plus, pour
C.
haitiensis,
s'ajoute l'importance nutritive de? characées, retrouvés principalement dans ,le secteur nord de l'Etang deMiragôane.
Par ailleurs, les zones d'eau libre reçoivent les individus
immatures et les femel~es en phase de maturation sexuelle post-fraie,
de l'espèce
S.
mossambiaus.
En résumé, il appert une utilisation intensive des zones
lacustres d'une profondeur inférieure à ~ mètres, couplée à un
couvert végétal très dense. où plus de 60% de cet habitat se retrouve
dans le secteur nord de l'Etang de Miragôane. le lac B Ces
terrasses localisent donc les contraintes écologiques de la
produc-tivité des Gichlidés de l,' Etang de Miragôane.
(
/ - ---~ ----~ 32. " / Chapitre 3 ~~
.
L'utilisation actuelle. des ressources de (
l'Etang de Miragôane, Haïti, qans la
vie rurale, et l'optimisation
possible de ce potentiel.
"
Soumis pour publication à la revue Journal of Applied Eco1ogy
/
Dernière étape de l'étude socio-écologique de l'Etang de
Miragôane, cette section détermine l'importance relative des
ressources inventoriées et analysées dans les deux chapitres
précédents. Les sujets élaborés identifient socialement le pêcheur
rural, le rendement de ses engins de pêche et le débouché de ses
captures dans les marchés locaux, via les revendeuses. Puis
l'analyse s'oriente vers la situation des éléocharides, qui
sup-portent un grappillage intense de la part du paysan; c'est-à-dire
qu'ils sont à la limite de la cueillette et de la culture proprement
dite. L~s points analysés définissen~ donc le profil social du 1
"coupeur de jonc", les revenus attribuables à cette ressource et
son rayonnement sur l'économie rurale.
En dernier lieu nous proposons des alternatives pour optimiser
l'utilisation des ressources aqu~tiques.
34.
3.1 INTRODUCTION
3.1.1 Les antécédents.
L'Etang de Miragôane, une des principales zones lacustres d'Haïti, attire depuis des décennies l'attention des officiels du pays et des coopérants étrangers, en tant que ressource sous-exploitée, soit en pêcheries (Lin 1952, Lovell et Moss 1971), soit en irrigation
(D.R.I.P.P. 1978a). Certains aménagements ont déjà été tentés avec plus ou moins de succès. Ainsi, en 1951 on introduisit un
-~----cichlidé africain,
Sarotherodon
m08sambicu8 Peters, afin de pallier à la faible productivité ichthyologique de ces eaux (Lin 1952).Aussi, au cours des années 50, sous l'égide américain-e, on construisit - une station de pompage présentement ~n ruine, sur le site de l'Etang
de Miragôane, afin d'irriguer les terres de la plaine adjacente.
Au cours de cette période, aucune analyse objective des resso~rces
disponibles et de leur apport économique en milieu rural n'a été considérée. En 1978, le gouvernement ha~tien en coopération avec l'Agence Canadienne de Développement International, décide de planifier un projet d'irrigation similaire à celui des américains, et ce durant la saison sèche. L'abaissement du niveau d'eau à
cette période de l'année, où il ac~use une cote minimale, pourrait éventuellement affecter les richesses aquatiques présentement disponibles.
l',
3.1.2 Les objectifs.
Cette étude vise d'abord à déterminer le rôle économique actuel
1
"
o
(
de l'Etang de Miragôane et ses implications dans le' monde rural connexe. Nous présentons donc ici le profil social des exploitants, une évaluati~n économique des ressources et leur rayonnement spatial. La compréhension des traits de la vie rurale contribuera à une
meilleure exploitation des ressources, tel que souligné par Moral (1961):
"c'est en partie pour n'en avoir pas même soupçonné l' exis,tence que bien des expé-riences de modernisation ont abouti à des échecs."
Pour terminer, nous développons diverses hypothèses concernant l'utilisation optimale des ressources'aquatiques disponibles.
3.1.3 Le site (figure 4).
La plaine marécageuse de l'Etang de Miragôane,.~à environ 7S km à l'ouest de Port-au-Prince, occupe 19.6 km2 de la section sud-haïtienne
d'Hispaniola, par 20°39' de latitude nord, 73°06' de longitude ouest. Elle réunit 3 lacs et elle draine un bassin versant de 149 km2
(cf. chapitre 2).
3.2 MATERIEL ET METHODES
3.2.1 Les recensements.
L'outil fondamental pour les survols sociologiques demeure le recensement. Celui accompli pour la ci~~onStance comporte une
l
section d'identification sociale du répo~dant, soient le sexe,
l
'.---...
l'âge, la zone d'origine, le statut civil, et le nombre d'enfants.