Associations entre les comportements alimentaires et
l’endocannabinoïdome
Mémoire
Gabrielle Rochefort
Maîtrise en nutrition - avec mémoire
Maître ès sciences (M. Sc.)
Associations entre les comportements alimentaires
et l’endocannabinoïdome
Mémoire
Gabrielle Rochefort
Sous la direction de :
Alain Veilleux, directeur de recherche
Véronique Provencher, codirectrice de recherche
Résumé
Mondialement, la prévalence de l’obésité ne cesse d’augmenter et certains traits du comportement alimentaire ont été identifiés comme facteurs de risque de cette condition. En parallèle, l’endocannabinoïdome, un système de signalisation lipidique, a été démontré comme jouant un rôle clé dans la régulation de l’homéostasie énergétique et de la prise alimentaire. Des évidences scientifiques suggèrent que des altérations dans ce système de signalisation sont impliquées dans le développement de l’obésité, mais également dans l’étiologie des troubles du comportement alimentaire. Toutefois, peu d’études relient l’endocannabinoïdome aux comportements alimentaires non pathologiques. L’objectif de ce projet de maitrise vise donc à étudier les associations entre les paramètres de l’endocannabinoïdome et certains comportements alimentaires non pathologiques tels que la restriction cognitive, la désinhibition, la susceptibilité à la faim et l’alimentation intuitive. Pour ce faire, les niveaux plasmatiques des médiateurs de l’endocannabinoïdome appartenant à la famille des N-acyl-éthanolamines et des 2-monoacyl-glycérols ont été mesurés dans une cohorte de 190 participants. Les comportements alimentaires ont été évalués à l’aide du Three-Factor Eating Questionnaire et de l’Intuitive Eating Scale-2. Les résultats de ce projet suggèrent que certains médiateurs de l’endocannabinoïdome dérivés d’acides gras polyinsaturés oméga-3 appartenant à la famille des 2-monoacyl-glycérols sont positivement associés à l’alimentation intuitive. À l’inverse, presque aucune association n’a été observée entre les médiateurs de l’endocannabinoïdome et la restriction cognitive, la désinhibition ainsi que la susceptibilité à la faim. Les résultats de la présente étude suggèrent un rôle potentiel de certains médiateurs de l’endocannabinoïdome dérivés d’acides gras polyinsaturés oméga-3 dans la régulation des comportements alimentaires sains et adaptatifs.
Abstract
Worldwide, the prevalence of obesity continues to increase, and some eating behavior traits have been identified as risk factors for this condition. In parallel, the endocannabinoidome, a lipid signaling system comprising several mediators, has been shown to play a critical role in the regulation of energy homeostasis and food intake. Scientific evidence suggests that alterations of the endocannabinoidome are implicated in obesity but also in the etiology of eating disorders. However, few studies have linked the endocannabinoidome to non-pathological eating behaviors. The objective of this master's project is therefore to study the associations between parameters of the endocannabinoidome and non-pathological eating behaviours such as cognitive restraint, disinhibition, susceptibility to hunger and intuitive eating. To do this, plasma levels of several endocannabinoidome mediators belonging to the N-acyl-ethanolamine and 2-monoacyl-glycerol families were measured in a cohort of 190 participants. Eating behaviours were assessed using the Three-Factor Eating Questionnaire and the Intuitive Eating Scale-2. The results of this project demonstrate that some endocannabinoidome mediators derived from omega-3 polyunsaturated fatty acids belonging to the 2-monoacyl-glycerol family are positively associated with intuitive eating. Conversely, almost no association was observed between the endocannabinoidome mediators and cognitive restraint, disinhibition and susceptibility to hunger. The results of the present study suggest a potential role of endocannabinoidome mediators derived from omega-3 polyunsaturated fatty acids in the regulation of healthy and adaptive eating behaviors.
Table des matières
Résumé ... ii
Abstract ... iii
Table des matières ... iv
Liste des figures ... vi
Liste des tableaux ... vii
Liste des abréviations ... viii
Avant-propos ... x Introduction ... 1 Chapitre 1 : Problématique ... 4 1.1 L’obésité ... 4 1.1.1 Définition de l’obésité ... 4 1.1.2 La prévalence de l’obésité ... 4
1.1.3 Les causes de l’obésité ... 5
1.1.4 Les conséquences de l’obésité ... 6
1.2 Les comportements alimentaires ... 6
1.2.1 L’étude des comportements alimentaires ... 6
1.2.2 Définitions et mesures ... 7
1.2.2 Restriction cognitive, désinhibition et susceptibilité à la faim ... 9
1.2.3 Alimentation intuitive ... 10
1.2.4 Les comportements alimentaires et le poids corporel ... 13
1.3 L’endocannabinoïdome ... 15
1.3.1 Définition du système endocannabinoïde et de l’endocannabinoïdome ... 15
1.3.2 Facteurs associés avec les médiateurs plasmatiques de l’endocannabinoïdome 18 1.3.3 L’endocannabinoïdome et l’homéostasie énergétique ... 20
1.3.4 L’endocannabinoïdome et la régulation de la prise alimentaire ... 22
1.3.5 L’endocannabinoïdome et les troubles du comportement alimentaire ... 24
1.3.6 Pertinence d’explorer les liens entre les comportements alimentaires et l’endocannabinoïdome ... 27
Chapitre 2 : Mise en contexte, objectifs et hypothèses ... 29
2.1 Mise en contexte ... 29
2.2 Objectifs ... 29
Chapitre 3 : L’alimentation intuitive est associée avec des niveaux circulants plus élevés de
médiateurs de l’endocannabinoïdome dérivés d’acides gras polyinsaturés oméga-3 ... 31
3.1 Résumé ... 33 3.2 Abstract ... 34 3.3 Introduction ... 35 3.4 Methods ... 37 3.4.1 Study population ... 37 3.4.2 Dietary intake ... 37 3.4.3 Anthropometric measurements ... 38
3.4.4 Circulating endocannabinoids and related mediators ... 38
3.4.5 Eating behaviours ... 39 3.4.6 Statistical analyses ... 39 3.5 Results ... 40 3.6 Discussion ... 42 3.7 References ... 48 3.8 Tables ... 54 3.9 Figures ... 60 Conclusion générale ... 62 Bibliographie ... 68
Liste des figures
Chapitre 1Figure 1. N-arachidonoyl-éthanolamine et 2-arachidonoyl-glycérol ... 16
Chapitre 3
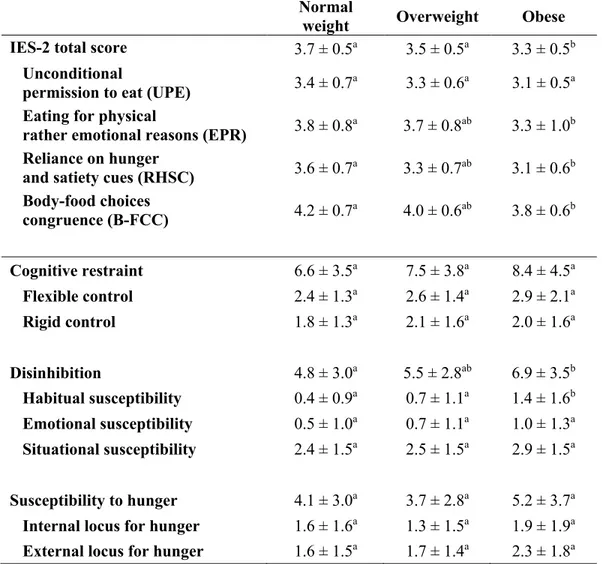
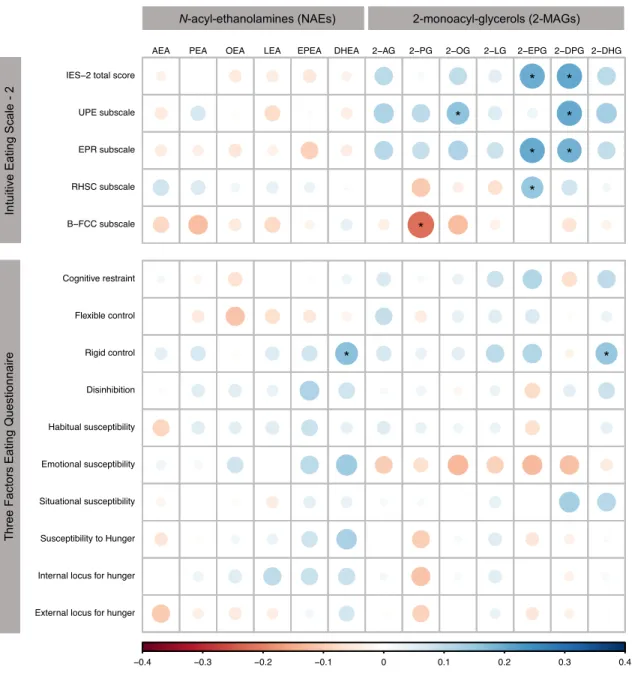
Figure 1. Associations of circulating NAEs and 2-MAGs with IES-2 and TFEQ scores. ... 60 Figure 2. IES-2 scores stratified by tertiles of circulating levels of 2-EPG and 2-DPG and by BMI categories. ... 61
Liste des tableaux
Chapitre 1Tableau 1. 10 principes de l’alimentation intuitive ... 11 Tableau 2. Congénères des N-acyl-éthanolamines et des 2-monoacyl-glycérols et leur acides gras précurseur ... 18
Chapitre 3
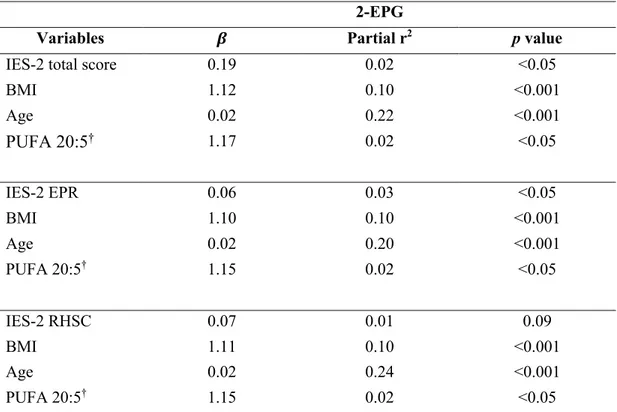
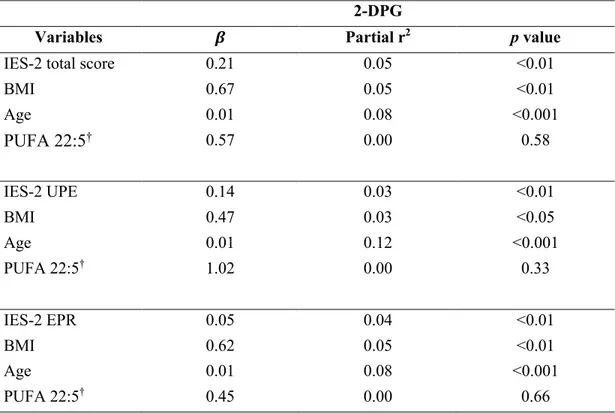
Table 1. Anthropometric characteristics of the participants. ... 54 Table 2. IES-2 and TFEQ total and subscale scores according to sex. ... 55 Table 3. IES-2 and TFEQ total and subscale scores according to BMI categories. ... 56 Table 4. Partial Spearman’s rank correlation coefficients between circulating 2-MAGs and IES-2 total and subscales scores independently of BMI and age. ... 57 Table 5. Multiple linear regression analysis for circulating levels of 2-EPG. ... 58 Table 6. Multiple linear regression analysis for circulating levels of 2-DPG. ... 59
Liste des abréviations
AEA: N-arachidonoyl-éthanolamine AGPI: Acide gras polyinsaturé ARN: Acide ribonucléique CB1: Cannabinoïde de type 1 CB2: Cannabinoïde de type 2
DHEA: N-docosahexaenoyl-éthanolamine EPEA:N-eicosapentaenoyl-éthanolamine FAAH: Fatty acid amide hydrolase 1
GPCR: Récepteur couplé à la protéine G (G protein‐coupled receptor) IES-2: Intuitive Eating Scale-2
IMC: Indice de masse corporelle LEA: N-linoleoyl-éthanolamine MAG: Monoacyl-glycerol lipase
NAAA: N-acylethanolamine acid amidase hydrolase NAE: N-acyl-éthanolamine
OEA: N-oleoyl-éthanolamine PEA: N-palmitoyl-éthanolamine
PPAR: Peroxisome proliferator-activated receptor TFEQ: Three-Factor Eating Questionnaire
TRPV: Transient receptor potential cation channel 2-AG: 2-arachidonoyl-glycérol 2-DHG: 2-docosahexaenoyl-glycérol 2-DPG:2-docosapentaenoyl-glycérol 2-EPG:2-eicosapentaenoyl-glycérol 2-LG: 2-linoleoyl-glycérol 2-MAG: 2-monoacyl-glycérol
2-OG: 2-oleoyl-glycérol 2-PG: 2-palmitoyl-glycérol
Avant-propos
J’ai fait mon entrée dans le monde de la recherche à la fin de ma première année de baccalauréat en nutrition. À cette époque, j’avais décidé de joindre l’équipe du Dr Alain Veilleux afin d’effectuer différents stages à titre d’auxiliaire de recherche en nutrition. La réalisation de ces stages a été un point culminant dans mon parcours universitaire en confirmant mon intérêt pour la recherche en nutrition et mon désir de poursuivre mes études universitaires au 2e cycle. J’ai donc débuté une maitrise en nutrition en janvier 2019 sous la
direction du Dr Alain Veilleux et la codirection de la Dre Véronique Provencher à l’Institut sur la nutrition et les aliments fonctionnels (INAF).
Au cours de ma maitrise, j’ai eu la chance de travailler avec les données du projet eMECA dont l’objectif principal était d’étudier les interactions de l'axe microbiome intestinal - endocannabinoïdome avec les facteurs environnementaux de l'hôte et l'état de santé intestinale, métabolique et mentale. À mon entrée à la maitrise, le recrutement et la collecte de données étaient terminés depuis octobre 2018. Toutefois, j’étais déjà bien familière avec le projet eMECA ayant eu l’occasion de participer au recrutement des participants et à la collecte de données lors de mon stage à l’été 2018. Dans le cadre de mon projet de maitrise, je me suis intéressée plus précisément aux associations entre l’endocannabinoïdome et les comportements alimentaires. J’ai donc eu la chance de travailler sur un projet de recherche combinant à la fois des aspects plus fondamentaux et psychologiques de la nutrition. J’ai effectué les analyses statistiques, l’interprétation des résultats, la présentation des résultats lors de congrès et réunions scientifiques ainsi que la rédaction d’un article scientifique à titre de première auteure. Cet article scientifique a été publié en septembre 2020 dans la revue Appetite et sera présenté dans le chapitre 3 du présent mémoire. À cet égard, je tiens à remercier tous les coauteurs de cet article pour leur collaboration et leur expertise, soit Véronique Provencher, Julie Perron, Sophie Castonguay-Paradis, Sébastien Lacroix, Cyril Martin, Nicolas Flamand, Vincenzo Di Marzo et Alain Veilleux.
La réalisation de mon projet de maitrise n’aurait pas pu être possible sans la contribution de nombreuses personnes. Tout d’abord, je tiens à remercier mon directeur de maitrise, Alain Veilleux, pour son aide précieuse. Tout au long de ma maitrise, il a fait preuve d’une grande
écoute et m’a transmis de nombreuses connaissances et qualités indispensables à la recherche. De plus, il a joué un rôle clé dans mon parcours universitaire et je le remercie grandement pour sa confiance, sa compréhension, son soutien, ses nombreux conseils et surtout, sa grande disponibilité. Je tiens également à remercier ma codirectrice, Véronique Provencher, pour le partage de son expertise ainsi que pour sa grande disponibilité tout au long de ma maitrise. Son expertise sur les comportements alimentaires a été essentielle à la réalisation de ce projet de recherche et sans sa contribution, je n’aurais pas été en mesure d’atteindre mes objectifs.
Je tiens également à remercier tous les membres de l’équipe Veilleux sans qui mon parcours à la maitrise n’aurait pas été aussi agréable et enrichissant. Entre autres, je remercie Julie Perron pour son soutien et ses conseils dans la réalisation de mon projet de maitrise. Je tiens également à remercier Sébastien Lacroix pour son aide précieuse dans la réalisation de nombreuses analyses statistiques et son esprit critique. Également, un merci spécial à Sophie Castonguay-Paradis avec qui j’ai eu la chance de travailler conjointement sur le projet eMECA pour son aide, ses conseils et sa grande écoute. De plus, je tiens à remercier tous les autres étudiants de l’équipe Veilleux, soit Bénédict Allam-Ndoul, Florent Pechereau, Frédéric Bégin ainsi que Briscia Tinoco. De belles amitiés se sont créées avec chacun d’entre vous rendant mon parcours à la maitrise mémorable.
Je considère également très important de remercier tous les membres de la Chaire d’excellence en recherche du Canada sur l’axe microbiome intestinal – endocannabinoïdome dans la santé métabolique. Plus précisément, je tiens à remercier Vincenzo Di Marzo pour sa grande expertise ainsi que Nicolas Flamand et Cyril Martin pour leur contribution à mon projet de maitrise.
Pour finir, un gros merci à tous mes ami(e)s et collègues de l’INAF. Je me considère très chanceuse d’avoir eu la chance de partager d’aussi beaux moments avec vous et d’avoir fait votre rencontre durant ma maitrise.
Introduction
Dans les dernières années, la prévalence de l’obésité et ses complications n’a cessé d’augmenter mondialement (Organisation mondiale de la santé, 2020a). Les études scientifiques ont mis en relief plusieurs facteurs impliqués dans l’étiologie de cette condition maintenant considérée comme une maladie par plusieurs organismes (Wharton et al., 2020). Entre autres, certains traits du comportement alimentaire ainsi qu’une suractivation de l’endocannabinoïdome ont été suggérés comme facteurs de risque de l’obésité (Chaput et al., 2009; Cristino et al., 2014).
Les comportements alimentaires font l’objet de plusieurs recherches depuis de nombreuses années. Les études s’intéressent notamment à mieux comprendre les facteurs associés aux comportements alimentaires et comment ces derniers influencent les choix alimentaires ainsi que le poids corporel des individus. Par exemple, plusieurs études ont révélé que les individus qui tentent de contrôler leur apport alimentaire dans le but de perdre ou de maintenir un certain poids corporel ont malgré tout un indice de masse corporelle (IMC) plus élevé et ne réussissent pas à éviter un gain de poids au fil du temps (Chaput et al., 2009; Lesdéma et al., 2012). De plus, la susceptibilité à la faim a été associée à l’embonpoint et l’obésité (Harden et al., 2009; Lesdéma et al., 2012), et il a été observé que la désinhibition alimentaire semble être un facteur de risque de l’obésité (Chaput et al., 2009). À l’inverse, certaines études ont démontré que les individus qui sont à l’écoute de leurs signaux physiologiques de faim et de satiété ont tendance à avoir un IMC plus faible en plus d’avoir une meilleure santé psychologique (Van Dyke & Drinkwater, 2014).
En parallèle, la littérature scientifique a démontré que l’endocannabinoïdome, un système de signalisation lipidique, joue un rôle clé dans le contrôle de l’homéostasie énergétique en favorisant le stockage d’énergie et en régulant la prise alimentaire. Ce système est largement distribué dans plusieurs organes clés du contrôle du métabolisme énergétique tels que le tissu adipeux, le foie et les muscles, et des altérations de ce système de signalisation ont été observées conjointement avec l’obésité suggérant une suractivation de l’endocannabinoïdome dans le développement de cette maladie (Silvestri & Di Marzo, 2013). L’endocannabinoïdome est également étroitement impliqué avec d’autres systèmes
de signalisation dans la régulation de la prise alimentaire. De nombreuses études ont démontré que les récepteurs de l’endocannabinoïdome sont distribués dans différentes régions du cerveau impliquées dans le contrôle de l’alimentation homéostatique (c.-à-d. basée sur la consommation de nourriture afin de rétablir la balance énergétique), mais également dans le contrôle de l’alimentation hédonique (c.-à-d. basé sur la consommation de nourriture pour le plaisir) (Di Marzo et al., 2009). De plus, des récepteurs de l’endocannabinoïdome sont également exprimés dans l’intestin et l’interaction de ces récepteurs avec certains médiateurs module la prise alimentaire (Sihag & Jones, 2018).
Certains groupes de chercheurs ont également souhaité mieux comprendre le rôle potentiel de l’endocannabinoïdome dans l’étiologie des troubles du comportement alimentaire. Entre autres, des altérations dans les niveaux plasmatiques de certains médiateurs de l’endocannabinoïdome ont été observées chez des femmes souffrant d’anorexie mentale et d’accès hyperphagiques (Monteleone et al., 2005). De plus, des changements dans les niveaux sanguins d’ARN messager d’un récepteur de l’endocannabinoïdome ont été observés chez les individus souffrant d’un trouble du comportement alimentaire (Frieling et al., 2009). Des études de génétiques ont également été conduites afin d’identifier des gènes de l’endocannabinoïdome impliqués dans l’étiologie des troubles du comportement alimentaire et des associations entre certains polymorphismes de gènes codant pour différents récepteurs ou enzymes de l’endocannabinoïdome et les troubles du comportement alimentaire ont été observées (Ishiguro et al., 2010; Monteleone et al., 2009).
Ces évidences scientifiques portent donc à croire que l’endocannabinoïdome pourrait influencer les traits du comportement alimentaire des individus. Dans le but de mieux comprendre le rôle potentiel de l’endocannabinoïdome dans la régulation des comportements alimentaires et les facteurs influençant ceux-ci, le projet de maitrise présenté dans le cadre de ce mémoire s’intéresse aux associations entre les médiateurs plasmatiques de l’endocannabinoïdome et certains traits du comportement alimentaire tels que la restriction cognitive, la désinhibition, la susceptibilité à la faim et l’alimentation intuitive.
Le présent mémoire est divisé en trois chapitres. Dans un premier temps, une brève revue de la littérature sur la problématique de l’obésité en association avec les comportements alimentaires et l’endocannabinoïdome est abordée. Dans le second chapitre, les objectifs spécifiques ainsi que les hypothèses de la présente recherche sont présentés. Le troisième chapitre de ce mémoire décrit la méthodologie ainsi que les résultats du présent projet sous la forme d’un article scientifique écrit en anglais et publié dans la revue Appetite. Pour finir, ce mémoire se termine par une discussion et une réflexion des principaux résultats de recherche et perspectives.
Chapitre 1 : Problématique
1.1 L’obésité
1.1.1 Définition de l’obésité
Selon l’Organisation mondiale de la santé, le surplus de poids et l’obésité sont définis comme «une accumulation anormale ou excessive de graisse qui présente un risque pour la santé» (Organisation mondiale de la santé, 2020b). À l’échelle populationnelle, l’IMC (poids (kg) /taille (m)2) est une mesure qui est utilisée afin de classifier l’obésité. L’obésité
est alors définie comme un IMC supérieur à 30 et classée en sous-catégories, soit l’obésité de classe 1 (IMC 30-34,9), de classe 2 (IMC 35-39,9) et de classe 3 (IMC 40 et plus). Plus l’IMC augmente, plus le risque de complications pour la santé associées à l’excès de graisse augmente. Cependant, à l’échelle des individus, cette mesure présente plusieurs limitations et ne permet pas de détecter adéquatement les complications associées à cette condition. D’un point de vue clinique, de nouvelles approches pour diagnostiquer et évaluer l’obésité ont été proposées et prennent en compte à la fois les paramètres métaboliques, physiques et psychologiques de cette condition (Wharton et al., 2020).
Plusieurs organismes, tels qu’Obésité Canada, l’Association médicale canadienne, l’American Medical Association et l’Organisation mondiale de la santé, considèrent désormais l’obésité comme une maladie chronique étant donné que sa gestion est un processus qui doit se faire à long terme (Obésité Canada, 2020b). Cette maladie complexe dont la prévalence ne cesse d’augmenter contribue à une augmentation du risque de complications médicales à long terme ainsi qu’à une réduction de l’espérance de vie (Wharton et al., 2020).
1.1.2 La prévalence de l’obésité
Depuis 1975, le nombre de cas d’obésité a presque triplé à l’échelle mondiale. Il est estimé qu’en 2016, plus de 1,9 milliard d’adultes dans le monde présentaient un surplus de poids et parmi ces individus, 650 millions souffraient d’obésité représentant 13% de la population adulte mondiale (Organisation mondiale de la santé, 2020a). Au Canada, selon l’Enquête sur
la santé dans les collectivités canadiennes de 2004, 23,1% des adultes canadiens souffraient d’obésité tandis qu’en 2015, se chiffre s’élevait à 26,7% (Gouvernement du Canada, 2017b). L’obésité est aujourd’hui considérée comme un important problème de santé publique et représente un lourd fardeau économique (Canada, 2011).
1.1.3 Les causes de l’obésité
L’obésité est le résultat d’un déséquilibre entre les calories consommées et dépensées. L’apport énergétique est alors supérieur à la dépense ce qui résulte, à long terme, à une balance énergétique positive. Une multitude de facteurs complexes tels que des facteurs génétiques, métaboliques, comportementaux et environnementaux interagissent entre eux et causent se déséquilibre énergétique (Obésité Canada, 2020a; Wharton et al., 2020). Par exemple, il est bien établi que la régulation de l’appétit est déterminée en partie par des interactions complexes au niveau du système nerveux central entre les composantes du système homéostatique, du système hédonique et du contrôle exécutif du lobe frontal. La communication entre ces systèmes est influencée par l’interaction de plusieurs médiateurs issus de différents tissus tels que le foie, le tissu adipeux et l’intestin. Il semblerait que l’interconnectivité de ces réseaux serait altérée dans l’obésité (Wharton et al., 2020). Des altérations dans le système endocannabinoïde ont, entre autres, été proposées comme étant l’une des causes sous-jacentes de l’hyperphagie et de l’obésité (Cristino et al., 2014).
Par ailleurs, une étude réalisée au Québec a mis en relief plusieurs facteurs de risque de l’embonpoint et de l’obésité chez l’adulte. Parmi ces facteurs, on retrouve un sommeil de courte durée, un faible apport alimentaire en calcium, l’absence d’activité physique à intensité élevée, la non-consommation de suppléments de vitamines et de minéraux, un apport alimentaire élevé en lipides et une consommation élevée d’alcool. De plus, les chercheurs de cette étude ont également démontré que certains traits du comportement alimentaire sont associés à l’embonpoint et l’obésité. Une désinhibition alimentaire, une susceptibilité à la faim ainsi qu’une restriction cognitive élevées constituent des facteurs de risque de l’embonpoint et de l’obésité chez l’adulte (Chaput et al., 2009).
En outre, il a été suggéré que des altérations dans la sensibilité aux signaux de faim et de satiété pourraient également être en cause dans le développement de l’obésité. En effet, certains individus souffrant d’obésité rapportent avoir de la difficulté à reconnaître leurs signaux de faim et/ou de satiété. Comparativement aux individus avec un poids normal, les sujets souffrant d’obésité sont caractérisés par des altérations dans leurs signaux de satiété (Drapeau et al., 2011). De plus, une faible réponse aux signaux de satiété a aussi été associée avec un IMC plus élevé chez l’adulte (Hunot et al., 2016). Il a également été suggéré qu’une sensibilité plus élevée à la récompense pourrait être associée à une surconsommation d’aliments et un IMC plus élevé (Drapeau et al., 2011).
1.1.4 Les conséquences de l’obésité
L’obésité peut avoir plusieurs conséquences sur la santé physique et psychologique. Cette condition affecte le bien-être et augmente le risque de développer plusieurs problèmes de santé tels que le diabète de type 2, l’hypertension artérielle, les maladies cardiovasculaires, les troubles musculo-squelettiques et plusieurs autres problèmes de santé. De plus, l’obésité a été associée à une augmentation du risque de développer certains cancers tel que le cancer du côlon, des reins, de l’œsophage, de l’endomètre et du sein (Obésité Canada, 2020b; Organisation mondiale de la santé, 2020a; Wharton et al., 2020).
Compte tenu des impacts pour la santé que présente l’obésité, il est important de bien comprendre les facteurs associés à cette condition ainsi que les interactions entre ceux-ci afin de mieux prévenir et traiter cette maladie. Dans les prochaines sections de ce mémoire, deux composantes associées à l’obésité, soit les comportements alimentaires ainsi que l’endocannabinoïdome, seront abordées plus en détail. De plus, les liens présumés entre ces deux composantes seront décrits.
1.2 Les comportements alimentaires
1.2.1 L’étude des comportements alimentaires
Les traits du comportement alimentaire des individus ont été associés avec plusieurs variables dans la littérature. Il a été démontré que les comportements alimentaires sont d’importants déterminants des choix alimentaires des individus et influencent de manière significative leur
apport alimentaire. Par exemple, certaines études ont observé que les individus restreignant leur alimentation tendent à surconsommer de la nourriture (Polivy & Herman, 1999) tandis que les mangeurs émotionnels ont tendance à consommer des aliments plus riches en sucre, en gras et en énergie pour faire face à des évènements stressants (Oliver et al., 2000). Il a été démontré que les individus ayant tendance à surconsommer des aliments ont un apport énergétique plus élevé (Lindroos et al., 1997) ainsi qu’une plus faible qualité de l’alimentation (Contento et al., 2005). À l’inverse, du point de vue de la saine alimentation, les femmes qui mangent en fonction de leurs signaux internes de faim et de satiété consomment plus de fruits et de légumes (Barad et al., 2019) et ont un apport énergétique plus faible (Camilleri et al., 2017).
Par ailleurs, certains traits du comportement alimentaire ont également été associés à de nombreuses reprises au poids corporel. Par exemple, il a été démontré que les individus avec une désinhibition alimentaire élevée ont un IMC plus élevé (Lindroos et al., 1997). De plus, différentes études ont également observé des associations entre les comportements alimentaires et certains indicateurs de santé physique et psychologique (Van Dyke & Drinkwater, 2014). Étant donné l’influence des comportements alimentaires sur les choix alimentaires des individus ainsi que sur différents indicateurs de santé, l’étude des comportements alimentaires suscite beaucoup d’intérêts. Dans la prochaine section de ce mémoire, certains traits du comportement alimentaire ainsi que leurs associations avec différentes variables, dont le poids corporel, seront abordés plus en détail.
1.2.2 Définitions et mesures
Selon l’Encyclopédie de la médecine comportementale, « le comportement alimentaire est un terme large qui englobe les choix et les motivations alimentaires, les habitudes alimentaires, les régimes et les problèmes liés à l’alimentation tels que l’obésité et les troubles du comportement alimentaire » (LaCaille, 2013). Le comportement alimentaire est très complexe et est déterminé par différents facteurs biologiques (p. ex. : la génétique, l’homéostasie énergétique), cognitifs (p. ex. : les croyances, les préférences alimentaires, les aversions, les émotions, l’influence des parents) et environnementaux ou externes (p. ex. : les facteurs sociaux, les facteurs économiques, la culture, la religion, l’éducation, l’accès et la
disponibilité des aliments) (Hernández Ruiz de Eguilaz et al., 2018). Bien que le terme comportement alimentaire soit très large, dans le cadre de ce mémoire il fait référence aux comportements alimentaires mesurés à l’aide de divers questionnaires tels que décrits dans les prochains paragraphes.
De nombreux outils ont été développés afin d’évaluer les comportements alimentaires des individus. D’une part, des questionnaires ont été développés afin d’identifier et d’évaluer certains symptômes ou certains traits associés aux comportements alimentaires non adaptatifs. Parmi les questionnaires évaluant les comportements alimentaires à risque, on retrouve l’Eating Disorder Inventory, conçu afin d’évaluer la psychopathologie des troubles du comportement alimentaire et les symptômes psychologiques associés (Garner et al., 1983), le Binge Eating Scale, évaluant les traits d’accès hyperphagiques des individus (Gormally et al., 1982), le Eating Attitude Test, identifiant la présence de troubles du comportement alimentaire (Garner et al., 1982) ainsi que le Emotional Eating Scale, mesurant la tendance des individus à manger en réponse à des émotions négatives (Arnow et al., 1995). D’autres questionnaires ont également été conçus afin de mesurer différentes dimensions du comportement alimentaire non pathologique. Le Three-Factor Eating Questionnaire (TFEQ) a été développé afin de mesurer trois dimensions de l’alimentation, soit la restriction cognitive, la désinhibition et la susceptibilité à la faim (Stunkard & Messick, 1985).
D’autre part, quelques questionnaires ont également été développés afin d’évaluer les comportements alimentaires d’un point de vue plus positif et permettent de mesurer les comportements alimentaires sains et adaptatifs. Parmi ces questionnaires, on retrouve le Mindful Eating Questionnaire (Framson et al., 2009), le eScatter Inventory (Lohse et al., 2007) ainsi que différentes échelles permettant de mesurer l’adhésion aux principes de l’alimentation intuitive telles que l’Intuitive Eating Scale-2 (IES-2) (Tylka & Kroon Van Diest, 2013).
Les différents outils présentés ci-dessus permettent donc d’évaluer différentes composantes du comportement alimentaire, chacun de ces outils présentant certaines forces et certaines
faiblesses. Dans le cadre de ce mémoire, les traits du comportement alimentaire qui seront abordés plus en détail sont la restriction cognitive, la désinhibition et la susceptibilité à la faim mesurées à l’aide du TFEQ ainsi que l’alimentation intuitive mesurée à l’aide l’IES-2. L’utilisation de ces deux questionnaires nous permet de couvrir un large spectre et de mesurer différentes dimensions du comportement alimentaire.
1.2.2 Restriction cognitive, désinhibition et susceptibilité à la faim
En 1985, Stunkard et Messick ont développé un questionnaire, le TFEQ, permettant de mesurer trois dimensions du comportement alimentaire. Ce questionnaire composé de 51 items permet de mesurer les niveaux de restriction cognitive (21 items), de désinhibition (16 items) et de susceptibilité à la faim (14 items) des individus (Stunkard & Messick, 1985).
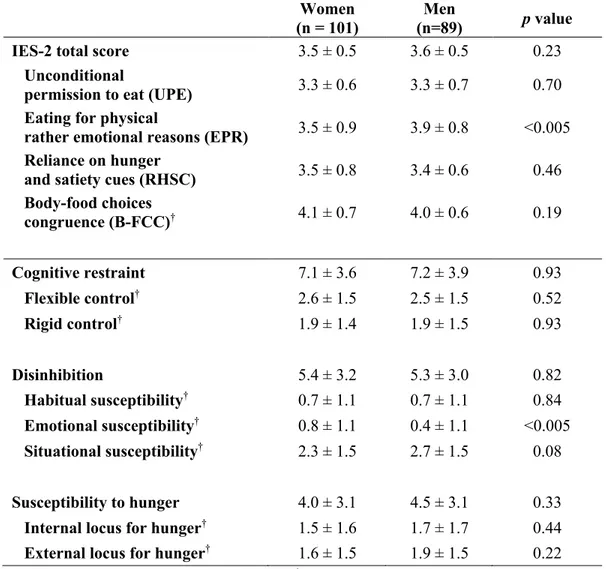
La restriction cognitive représente l’intention des individus de contrôler leur apport alimentaire dans le but de perdre du poids ou de maintenir un certain poids corporel (Stunkard & Messick, 1985; Westenhoefer et al., 1999). La restriction cognitive se divise en deux sous-échelles, soit la restriction rigide et la restriction flexible. La restriction rigide correspond à une approche dichotomique de l’alimentation, des régimes et du poids dans laquelle les aliments palatables sont interdits. La sous-échelle de la restriction flexible représente une approche plus graduée de l’alimentation, des régimes et du poids. Dans cette approche, les aliments palatables qui sont plus riches en calories sont permis en petite quantité, et ce, sans sentiment de culpabilité (Westenhoefer et al., 1999). Il a été observé que la restriction cognitive est plus élevée chez les femmes que chez les hommes (Lesdéma et al., 2012; Löffler et al., 2015; Provencher et al., 2003) et tend à augmenter avec l’âge (Drapeau et al., 2003; Löffler et al., 2015). De plus, bien que la restriction cognitive ait été associée à un apport énergétique plus faible (Provencher et al., 2003), une restriction cognitive élevée a aussi été associée à une surconsommation d’aliments dans des situations stressantes (Wardle et al., 2000). Il a également été observé que les individus avec une restriction rigide élevée ont des épisodes plus sévères et plus fréquents d’accès hyperphagiques. À l’opposé, la restriction flexible a été associée à des épisodes moins fréquents et moins sévères d’accès hyperphagiques ainsi qu’à un apport énergétique plus faible (Westenhoefer et al., 1999).
Ensuite, la désinhibition représente la tendance à la surconsommation associée à une perte de contrôle dans les aliments (Bond et al., 2001; Stunkard & Messick, 1985). La désinhibition peut être divisée en trois sous-dimensions, soit la susceptibilité habituelle, émotionnelle et situationnelle à la désinhibition. La susceptibilité habituelle à la désinhibition décrit les comportements qui peuvent survenir lorsque les circonstances prédisposent à une désinhibition récurrente. Les sous-dimensions de la susceptibilité émotionnelle et situationnelle à la désinhibition représentent, respectivement, la susceptibilité de surconsommer de la nourriture en réponse à des états affectifs négatifs (p. ex. : tristesse, anxiété, solitude) ou à des signaux environnementaux précis (p. ex. : la présence d’un individu qui surconsomme de la nourriture) (Bond et al., 2001). Il a été observé que les femmes ont une susceptibilité à la désinhibition plus élevée que les hommes (Drapeau et al., 2003; Lesdéma et al., 2012; Löffler et al., 2015). De plus, une susceptibilité à la désinhibition élevée a été associée à un apport énergétique plus élevé (Lindros et al.), des variations pondérales plus fréquentes, des épisodes plus nombreux de surconsommation alimentaire, certains désordres alimentaires et des niveaux plus faibles d’activité physique (Lowson et al. 1995).
Enfin, la susceptibilité à la faim représente la susceptibilité des individus à manger en réponse à la sensation ou à la perception de la faim (Bond et al., 2001; Stunkard & Messick, 1985). Deux sous-échelles découlent de cette dimension soit la faim interne et la faim externe. La faim interne représente une faim qui est interprétée et régulée de manière interne tandis que la faim externe correspond à une faim qui est déclenchée par des signaux externes (Bond et al., 2001). Une susceptibilité à la faim élevée a été associée, entre autres, à un IMC et un apport énergétique plus élevé (Provencher et al., 2003). De plus, il a été observé que les individus plus jeunes ont une susceptibilité à la faim plus élevée que les individus plus âgés (Harden et al., 2009; Löffler et al., 2015).
1.2.3 Alimentation intuitive
L’alimentation intuitive est une approche positive de l’alimentation basée sur l’écoute de ses signaux de faim et de satiété, plutôt que sur des signaux externes et émotionnels, afin de guider la prise alimentaire, et déterminer quand, quoi et quelle quantité d’aliments manger
(Tribole & Resch, 1995). Selon cette approche, le corps est parfaitement outillé pour déterminer la quantité ainsi que le type d’aliments à consommer afin de maintenir une bonne santé nutritionnelle ainsi qu’un poids corporel adéquat. Aucun aliment n’est interdit et aucune restriction nutritionnelle n’est imposée, à moins de souffrir de certaines maladies. Selon le principe de l’alimentation intuitive, le corps optera d’emblée pour une variété d’aliments offrant un équilibre nutritionnel (Van Dyke & Drinkwater, 2014). Dans cette approche, en plus d’entretenir une relation saine avec les aliments, la pensée et le corps, les émotions ainsi que le plaisir associés à la prise alimentaire sont pris en considération (Tribole & Resch, 1995). Le concept de l’alimentation intuitive est constitué de 10 principes qui ont été préalablement proposés par Tribole et Resch (Tribole et Resch 2012).
Tableau 1. 10 principes de l’alimentation intuitive
1. Rejeter la mentalité des diètes amaigrissantes qui crée de faux espoirs. 2. Honorer sa faim lorsque celle-ci est modérée.
3. Se permettre de manger tous les aliments souhaités.
4. Cesser de catégoriser les aliments comme étant «bons» ou «mauvais». 5. Considérer sa sensation de satiété pour savoir quand cesser de manger. 6. Découvrir la satisfaction et le plaisir de manger.
7. Vivre ses émotions sans nécessairement utiliser la nourriture. 8. Respecter son corps tel qu’il est au moment présent.
9. Être physiquement actif par plaisir et non pour perdre du poids. 10. Honorer sa santé et ses papilles gustatives par ses aliments préférés.
Note : Ces principes ont été tirés de la thèse de doctorat de Karine Gravel (Gravel, 2013; Tribole & Resch, 2012).
Afin de mesurer le concept de l’alimentation intuitive selon les principes élaborés par Tribole et Resch, l’IES-2 a été développée (Tylka & Kroon Van Diest, 2013). Cette échelle, qui est une version améliorée de l’Intuitive Eating Scale (Tylka, 2006), est composée de 23 items et permet de mesurer le degré d’adhésion aux principes de l’alimentation intuitive. Elle se divise en 4 sous-échelles soit la Permission inconditionnelle de manger, Manger pour des raisons
physiques plutôt qu’émotionnelles, la Référence aux signaux de faim et de satiété et la Congruence des Choix Aliments-Corps (Tylka & Kroon Van Diest, 2013).
Premièrement, la sous-échelle de la Permission inconditionnelle de manger évalue la volonté des individus à manger lorsque ceux-ci ont physiologiquement faim et à manger la nourriture souhaitée à ce moment. Les individus qui se permettent de manger de manière inconditionnelle n’étiquettent pas les aliments comme étant « bons » ou « mauvais » et l’absence d’aliments interdits rendrait ces individus moins susceptibles de surconsommer de la nourriture, d’avoir des accès hyperphagiques, ou encore, de se sentir coupables en mangeant. Deuxièmement, la sous-échelle Manger pour des raisons physiques plutôt qu’émotionnelles mesure la tendance des individus à manger pour satisfaire une faim interne plutôt que pour faire face à leurs émotions telles que l’anxiété, la solitude et l’ennui. La surconsommation d’aliments ainsi que les préoccupations alimentaires seraient plus faibles chez les femmes qui mangent pour des raisons physiques plutôt que pour faire face à leurs émotions. Troisièmement, la Référence aux signaux de faim et de satiété représente le degré de conscience et de confiance dans ses signaux internes de faim et de satiété pour guider ses comportements alimentaires. Les individus qui ne sont pas à l’écoute de leurs signaux physiologiques de faim et de satiété auraient plus tendance à se restreindre, à manger selon leurs émotions et à prendre du poids. Pour finir, la sous-échelle de la Congruence des Choix Aliments-Corps permet de mesurer le principe de la « l’alimentation bienveillante ». Ce principe reflète la tendance des individus à choisir des aliments honorant leur santé ainsi que leurs papilles gustatives. Cette sous-échelle mesure donc la capacité et le désir des individus à choisir des aliments nutritifs qui offrent énergie et vigueur au corps. Les individus avec une congruence des choix aliments-corps élevée ont tendance à choisir des aliments sains parce que c’est ce dont le corps a besoin et non parce qu’ils ressentent une certaine pression à agir de cette façon (Tylka & Kroon Van Diest, 2013).
Dans une revue systématique réalisée par Bruce et Ricciardelli, l’alimentation intuitive a été associée à une plus faible prévalence de troubles du comportement alimentaire, une meilleure image corporelle ainsi qu’un meilleur fonctionnement émotif chez les femmes (Bruce & Ricciardelli, 2016). De plus, une revue de la littérature réalisée par Van Dyke et Drinkwater
a démontré que l’alimentation intuitive est fortement associée à une meilleure santé psychologique (Van Dyke & Drinkwater, 2014). À l’inverse, les évidences reliant l’alimentation intuitive et la qualité de l’alimentation sont moins claires (Barad et al., 2019; Camilleri et al., 2017; Horwath et al., 2019). En effet, certaines études ont démontré que l’alimentation intuitive était positivement associée avec la consommation de fruits et de légumes (Barad et al., 2019; Madden et al., 2012) tandis que d’autres études n’ont observé aucune association (Horwath et al., 2019). De plus, certaines sous-échelles de l’IES-2 ont été associées avec d’autres aspects de l’alimentation tels que l’apport énergétique, la consommation de sucrerie et la qualité de la diète, mais plus d’études sont nécessaires afin d’éclaircir ces associations (Barad et al., 2019; Camilleri et al., 2017; Horwath et al., 2019). Il a également été observé qu’une intervention portant sur l’alimentation intuitive améliore la qualité de l’alimentation à court terme (Carbonneau et al., 2017). De plus, plusieurs études ont démontré que les mangeurs intuitifs ont un IMC plus faible que les mangeurs non intuitifs (Van Dyke & Drinkwater, 2014).
1.2.4 Les comportements alimentaires et le poids corporel
Les traits du comportement alimentaire des individus ont été associés à plusieurs reprises avec le poids corporel. Certaines études ont démontré que la restriction cognitive était négativement associé à l’IMC (Boschi et al., 2001; Foster et al., 1998). Entre autres, une augmentation plus importante de la restriction cognitive a été associée à une perte de poids plus importante durant un programme de perte de poids (Foster et al., 1998). À l’inverse, certaines études ont observé des associations positives entre la restriction cognitive et l’IMC (Chaput et al., 2009; Lesdéma et al., 2012). Il a été suggéré que les adultes qui restreignent leur apport alimentaire sont plus susceptibles d’avoir un gain de poids et de développer une obésité au fil du temps (Chaput et al., 2009). Chez les femmes, une restriction cognitive élevée semble également promouvoir le gain de poids tandis que chez les hommes la restriction cognitive est associée à une perte de poids (Drapeau et al., 2003). Une restriction rigide élevée a été associée à un IMC, un tour de taille et des niveaux de graisse corporelle plus élevés tandis que la restriction flexible a été négativement associée à l’IMC, la graisse corporelle, le tour de taille ainsi que les niveaux de tissus adipeux viscéral et sous-cutané (Provencher et al., 2003; Westenhoefer et al., 1999).
Par ailleurs, une désinhibition alimentaire et une susceptibilité à la faim élevées semblent être contreproductives afin de perdre du poids. Comparativement à la restriction cognitive, ces deux dimensions du comportement alimentaire ont été associées positivement à l’IMC de façon plus constante dans les études. En effet, la désinhibition alimentaire et la susceptibilité à la faim ont été associées à plusieurs reprises avec l’embonpoint et l’obésité tant chez les hommes que chez les femmes (Chaput et al., 2009; Harden et al., 2009; Lesdéma et al., 2012; Lindroos et al., 1997; Provencher et al., 2003). Parmi les trois dimensions du comportement alimentaire mesurées par le TFEQ, la désinhibition est la dimension qui est la plus fortement associée à l’obésité et un IMC élevé (Lesdéma et al., 2012; Lindroos et al., 1997). Une désinhibition alimentaire élevée promeut le gain de poids et augmente le risque de développer un embonpoint et une obésité (Chaput et al., 2009; Hays et al., 2002). De plus, la susceptibilité génétique à l’obésité, déterminée à l’aide de 97 polymorphismes nucléotidiques simples, semble être partiellement médiée par la désinhibition et la susceptibilité à la faim chez l’adulte (Jacob et al., 2018).
À l’opposé, étant donné que le principe de l’alimentation intuitive consiste à manger en fonction de ses signaux de faim et de satiété et que la prise alimentaire n’est pas déterminée par ses émotions ou par des signaux externes, manger de façon intuitive devrait être associé à un poids corporel ou un IMC plus faible. Selon une revue de la littérature, les individus avec une alimentation intuitive ont un IMC plus faible comparativement à des mangeurs non intuitifs. Ces résultats s’appliquent entre autres auprès des femmes et des étudiants universitaires (Van Dyke & Drinkwater, 2014). De plus, bien que l’objectif d’adopter une alimentation plus intuitive ne soit pas de perdre du poids, l’effet de l’implantation d’un programme basé sur l’alimentation intuitive sur le poids corporel a été étudié. Toujours selon cette revue de la littérature, il a été démontré que, bien que la participation à un tel programme n’entraine pas de perte de poids, ce type d’approche aiderait au maintien du poids corporel chez des femmes caucasiennes souffrant d’embonpoint ou d’obésité (Van Dyke & Drinkwater, 2014). Ainsi, comparativement aux approches traditionnelles des diètes qui à court terme mènent à une perte de poids suivie d’une reprise du poids corporel, l’adoption
d’une alimentation plus intuitive pourrait aider au maintien du poids corporel bien que d’autres études sur le sujet soient nécessaires.
En somme, comme démontré, les comportements alimentaires sont fortement associés avec le poids corporel et certaines dimensions du comportement alimentaire ont été associées à plusieurs reprises à l’embonpoint et l’obésité. L’utilisation du TFEQ et de l’IES-2 nous permet d’avoir une vue d’ensemble du spectre du comportement alimentaire et ainsi de bien étudier le lien entre les comportements alimentaires et différentes composantes biologiques. Dans la prochaine section de ce mémoire, l’endocannabinoïdome, un système de signalisation lipidique impliqué dans la régulation de l’homéostasie énergétique et qui a notamment été associé avec le poids corporel et les troubles du comportement alimentaire, sera abordé en détail. Les associations entre ce système de signalisation lipidique et certains traits du comportement alimentaire (c.-à-d. restriction cognitive, désinhibition, susceptibilité à la faim et alimentation intuitive) seront ensuite discutées.
1.3 L’endocannabinoïdome
1.3.1 Définition
du
système
endocannabinoïde
et
de
l’endocannabinoïdome
Dans les années 1950 et 1960, les principaux constituants du Cannabis Sativa, le cannabidiol ainsi que le D9-tetra-hydrocannabinol, ont été découverts. Quelques années plus tard, les deux
récepteurs couplés à la protéine G (GPCRs) via lesquels la molécule de D9-tetra-hydrocannabinol agit dans le corps ont été identifiés, soit le récepteur
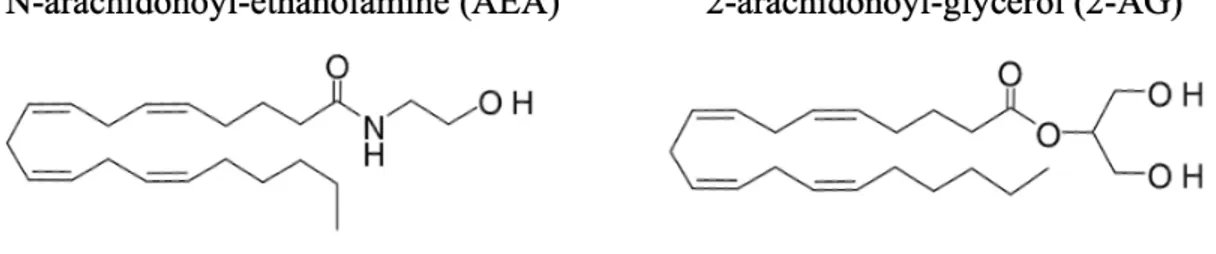
cannabinoïde de type 1 (CB1) et le récepteur cannabinoïde de type 2 (CB2). L’identification de ces deux récepteurs a mené à la caractérisation de leurs ligands endogènes, également appelé endocannabinoïdes, soit le N-arachidonoyl-éthanolamine (anandamide; AEA) ainsi que le 2-arachidonoyl-glycérol (2-AG). Le système endocannabinoïde a alors été désigné comme l’ensemble formé par les deux endocannabinoïdes (c.-à-d. AEA et 2-AG), leurs récepteurs CB1 et CB2 ainsi que leurs 5 enzymes principales de synthèse et de dégradation (Di Marzo, 2018).
Les endocannabinoïdes sont des médiateurs lipidiques dérivés d’acides gras polyinsaturés (AGPI) qui sont synthétisés à partir de précurseurs contenus dans les phospholipides des membranes cellulaires. L’AEA et le 2-AG sont constitués d’un acide gras oméga-6 arachidonique et d’une molécule d’éthanolamine ou de glycérol respectivement (Figure 1). Les niveaux de ces médiateurs lipidiques sont établis sur demande par des enzymes de synthèse (c.-à-d. N-acyl-phosphatidylethanolamine-hydrolysing phospholipase D, sn-1-specific diacylglycerol lipase-α, sn-1-specific diacylglycerol lipase-b) et de dégradation (c.-à-d. fatty acid amide hydrolase 1 (FAAH), monoacyl-glycerol lipase (MAGL)) (Di Marzo, 2018). Contrairement à certains médiateurs ou neurotransmetteurs, les endocannabinoïdes ne sont pas entreposés dans des vésicules après leur synthèse. Ces médiateurs sont immédiatement libérés afin d’agir à des moments et à des endroits précis dans le corps. Les endocannabinoïdes peuvent se lier aux récepteurs CB1 et CB2 de manière autocrine ou paracrine et leurs interactions avec ces récepteurs entrainent une multitude de réponses physiologiques liées au métabolisme énergétique, à l’inflammation ainsi qu’à l’immunité (Gatta-Cherifi & Cota, 2016). Les endocannabinoïdes sont présents dans le plasma ainsi que dans de nombreux tissus, notamment dans le tissu adipeux, le muscle, le foie et le cerveau (Silvestri & Di Marzo, 2013).
Les récepteurs CB1 et CB2 sont largement distribués dans le corps et permettent la régulation de plusieurs processus biochimiques. D’une part, les récepteurs CB1 sont principalement retrouvés au niveau des cellules du système nerveux central qui participent à la régulation de l’appétit, de la dépense énergétique et de la récompense. Également, les récepteurs CB1 sont exprimés dans différents tissus périphériques qui jouent un rôle clé dans le contrôle de l’homéostasie métabolique tels que le tissu adipeux, le foie, la pancréas et les muscles (Silvestri & Di Marzo, 2013). Certaines études précliniques ont démontré que l’activation
des récepteurs CB1 peut s’avérer positive dans le traitement de certaines maladies telles que les maladies neurologiques, les troubles neuropsychiatriques et les maladies inflammatoires de l’intestin (Di Marzo, 2018). D’autre part, les récepteurs CB2 sont principalement retrouvés dans les cellules et les tissus du système immunitaire. Ces récepteurs participent à la régulation de l’immunité ainsi qu’à la réponse inflammatoire. Les récepteurs CB2 sont également présents au niveau du système nerveux central, mais en moins grande quantité comparativement aux récepteurs CB1 (Silvestri & Di Marzo, 2013). De plus, bien que les endocannabinoïdes se fixent aux récepteurs CB1 et CB2, ces médiateurs lipidiques peuvent également interagir avec d’autres récepteurs cellulaires, résultant parfois à des effets opposés selon les récepteurs avec lesquels ces molécules interagissent. Par exemple, ces médiateurs se lient aux canaux ioniques transient receptor potentiel vanilloid (TRPVs) ainsi qu’aux récepteurs nucléaires peroxisome proliferator-activated receptors (PPARs) (Di Marzo, 2018).
L’AEA et le 2-AG font partie de deux grandes familles de lipides soit les N-acyl-éthanolamines (NAEs) et les 2-monoacyl-glycérols (2-MAGs) respectivement. Comme les endocannabinoïdes, les différents médiateurs de la famille des NAEs et des 2-MAGs, également appelés congénère des endocannabinoïdes, sont synthétisés à partir d’acides gras et sont formés soit d’une molécule d’éthanolamine ou d’une molécule de glycérol (tableau 2) (Di Marzo, 2018). Ces médiateurs sont synthétisés et inactivés par les mêmes enzymes que les endocannabinoïdes (Silvestri & Di Marzo, 2013). Toutefois, comparativement à l’AEA et au 2-AG, les congénères des endocannabinoïdes interagissent avec des récepteurs non-cannabinoïdes (c.-à-d. autres que les récepteurs CB1 et CB2) tels que les canaux ioniques TRPVs (p. ex. : TRPV1), les récepteurs nucléaires PPARs (p. ex. : PPARa, PPARg) et d’autres GPCRs (p. ex. : GPR55, GPR119). Une expansion du système endocannabinoïde a donc été proposée et est communément appelée l’endocannabinoïdome. Cette expansion est formée, en plus des molécules composant le système endocannabinoïde, des NAEs, des 2-MAGs, de certains médiateurs lipidiques possédant d’autres groupements amines ainsi que des récepteurs et des enzymes de conversions spécifiques à ces molécules (Di Marzo, 2018). Bien que ces médiateurs lipidiques, enzymes et récepteurs appartiennent à l’endocannabinoïdome, ces molécules ont
été, jusqu’à présent, moins étudiées que les molécules composant le système endocannabinoïde, ce qui explique pourquoi dans les prochaines sections de ce mémoire l’AEA, le 2-AG ainsi que les récepteurs CB1 et CB2 seront abordés plus en détail.
Tableau 2. Congénères des N-acyl-éthanolamines et des 2-monoacyl-glycérols et leur
acide gras précurseur
N-acyl-éthanolamines
(NAEs)
2-monoacyl-glycérols (2-MAGs)
Acides gras précurseur
N-arachidonoyl-éthanolamine (AEA)* 2-arachidonoyl-glycérol (2-AG)* Acide arachidonique (20 :4) N-palmitoyl-éthanolamine
(PEA) 2-palmitoyl-glycérol (2-PG) Acide palmitique (16 :0)
N-oleoyl-éthanolamine (OEA) 2-oleoyl-glycérol (2-OG) Acide oléique (18 :1) N-linoleoyl-éthanolamine (LEA) 2-linoleoyl-glycérol (2-LG) Acide linoléique (18 :2) N-eicosapentaenoyl-éthanolamine (EPEA) 2-eicosapentaenoyl-glycérol (2-EPG) Acide eicosapentaénoïque (20 :5) N-docosapentaenoyl-éthanolamine (DPEA) 2-docosapentaenoyl-glycérol (2-DPG) Acide docosapentaénoïque (22 :5) N-docosahexaenoyl-éthanolamine (DHEA) 2-docosahexaenoyl-glycérol (2-DHG) Acide docosahexaénoïque (22 :6) * Endocannabinoïdes
1.3.2 Facteurs associés avec les médiateurs plasmatiques de
l’endocannabinoïdome
Chez l’humain, la mesure des niveaux plasmatiques des médiateurs de l’endocannabinoïdome est fréquemment utilisée dans les études puisque cette mesure est beaucoup plus accessible. En effet, cette mesure est beaucoup moins invasive et beaucoup plus facile à récolter que, par exemple, la mesure des niveaux de ces médiateurs dans le tissu adipeux ou le muscle puisqu’elle nécessite qu’une prise de sang. Par contre, cette méthode comporte certaines faiblesses étant donné que le rôle exact des médiateurs de l’endocannabinoïdome en circulation ainsi que leur provenance (p. ex. : système nerveux central, tissu adipeux, muscle, intestin) ne sont pas bien connus. Les études portant sur l’endocannabinoïdome ont démontré que plusieurs facteurs semblent influencer les niveaux circulants de ces médiateurs. Parmi ces facteurs, on retrouve l’adiposité, les apports alimentaires, la pratique d’activité physique ainsi que les bactéries composant le microbiote
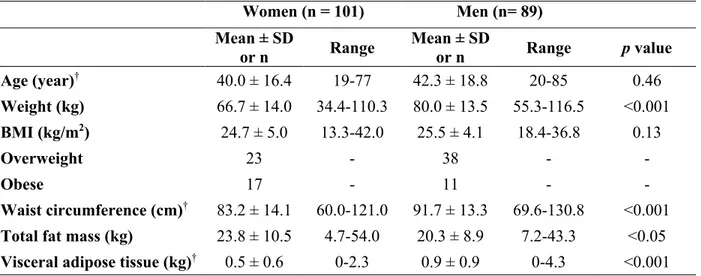
intestinal. En effet, les niveaux circulants des endocannabinoïdes, soit l’AEA et le 2-AG, ont été positivement associés avec différentes mesures d’adiposité et mesures anthropométriques telles que la graisse corporelle, le tissu adipeux viscéral, l’IMC et la circonférence de taille (Blüher et al., 2006a; Côté et al., 2007a; Engeli et al., 2005; Fanelli et al., 2018). Il a été démontré que les mesures de composition corporelle et de distribution des graisses corrèlent avec les niveaux plasmatiques des médiateurs de l’endocannabinoïdome. Plus précisément, la plupart des congénères des NAEs corrèlent positivement avec la masse grasse tandis que les congénères des 2-MAGs corrèlent positivement avec les niveaux de graisse viscérale (Castonguay-Paradis et al., 2020).
Les apports alimentaires ont également été associés avec les niveaux circulants des médiateurs de l’endocannabinoïdome. Plus précisément, il a été démontré que les niveaux circulants de certains congénères de NAEs sont altérés en réponse à une supplémentation en huiles riches en différents types d’acides gras et corrèlent positivement avec les niveaux plasmatiques de leur acide gras précurseur (Pu et al., 2016). De plus, selon une récente étude, les niveaux circulants de certains congénères des NAEs et des 2-MAGs sont associés avec l’apport alimentaire rapporté en acides gras précurseurs de ces congénères. Également, il a été observé que la consommation d’une diète méditerranéenne riche en acide gras oléique, eicosapentaénoïque et docosahexaénoïque entraine une augmentation des niveaux circulants de certains congénères des NAEs et des 2-MAGs (Castonguay-Paradis et al., 2020).
Par ailleurs, la pratique d’activité physique semble activer l’endocannabinoïdome et modifier les niveaux circulants de certains médiateurs. En effet, les niveaux circulants d’AEA et d’autres congénères des NAEs augmentent suivant la pratique d’activité physique à intensité modérée (Hillard, 2018). Il a été observé que la pratique d’activité physique d’intensité modérée à élevée est associée avec des niveaux circulants plus élevés d’AEA chez des femmes de poids normal et souffrant d’obésité (Fernández-Aranda et al., 2014). Bien que les effets de la pratique d’activité physique sur les niveaux de 2-AG soient moins évidents, une récente étude a démontré que la pratique d’activité physique est associée avec des niveaux circulants plus élevés d’AEA, mais également de 2-AG (Brellenthin et al., 2017).
Certaines études se sont également intéressées aux associations entre les médiateurs de l’endocannabinoïdome et le microbiote intestinal et suggèrent une association bidirectionnelle entre ces deux systèmes dans l’axe microbiome intestinal - endocannabinoïdome. En effet, l’endocannabinoïdome et le microbiote intestinal seraient interreliés et interviendraient dans la régulation du métabolisme (Cani et al., 2016). Il a été démontré que chez la souris, des changements au niveau de la composition du microbiote intestinal sont accompagnés par des changements dans l’endocannabinoïdome au niveau de l’intestin (Guida et al., 2018; Manca et al., 2020). L’endocannabinoïdome a également été associé avec la composition du microbiote intestinal dans la dysbiose associée à l’obésité (Lacroix et al., 2019). Par ailleurs, chez l’humain, l’abondance relative de certaines familles bactériennes corrèle avec les niveaux plasmatiques de certains congénères de la famille des NAEs et des 2-MAGs indépendamment de l’adiposité (Castonguay-Paradis et al., 2020).
Au cours des dernières années, la compréhension de l’endocannabinoïdome s’est grandement améliorée et plusieurs rôles physiologiques lui ont été attribués dont un rôle clé dans la régulation de l’homéostasie énergétique et de la prise alimentaire (Silvestri & Di Marzo, 2013). De plus, des altérations dans la signalisation de l’endocannabinoïdome ont également été associées avec différentes pathologies reliées à l’homéostasie énergétique telles que l’obésité, le diabète de type 2 ainsi que les troubles du comportement alimentaire et ce système a été la cible dans le traitement de plusieurs maladies (Di Marzo, 2018; Marco et al., 2012).
1.3.3 L’endocannabinoïdome et l’homéostasie énergétique
L’endocannabinoïdome joue un rôle clé dans la régulation de l’homéostasie énergétique, entre autres, en augmentant la prise alimentaire et en favorisant le stockage d’énergie. Sous certaines conditions, ce système intervient avec d’autres systèmes de signalisation homéostatique afin de restaurer l’homéostasie locale qui a été perturbée, et ce, en favorisant la production de médiateurs lipidiques (Di Marzo, 2008). Les endocannabinoïdes régulent la balance énergétique par leurs interactions avec les récepteurs cannabinoïdes exprimés au niveau du système nerveux central et dans différents tissus et organes périphériques (Li et al., 2011).
Plus précisément, l’endocannabinoïdome joue un rôle important dans plusieurs organes clés qui interviennent dans le contrôle de l’homéostasie énergétique tels que le tissu adipeux, le muscle, le foie ainsi que le pancréas (Gatta-Cherifi & Cota, 2016). Les récepteurs CB1 ainsi que différentes enzymes de synthèse et de dégradation des endocannabinoïdes sont exprimés au niveau des adipocytes blancs (Silvestri & Di Marzo, 2013). L’activation des récepteurs CB1 stimule l’adipogenèse ainsi la lipogenèse tandis que, à l’opposé, la stimulation du canal ionique TRPV1 par l’AEA inhiberait la lipogenèse (Di Marzo, 2008). Les hépatocytes ainsi que les cellules musculaires produisent des endocannabinoïdes et expriment également les récepteurs CB1. L’activation de ces récepteurs au niveau du foie entraine une accumulation de lipides tandis que ce système semble intervenir de manière négative sur les voies oxydatives au niveau du muscle (Silvestri & Di Marzo, 2013). Les récepteurs CB1 sont également exprimés au niveau des ilets pancréatiques et participent à la signalisation et à la libération de l’insuline (Gatta-Cherifi & Cota, 2016). L’activation in vitro des récepteurs CB1 augmenterait la sécrétion d’insuline (Silvestri & Di Marzo, 2013). Par ailleurs, la stimulation des récepteurs GPR119 par l’OEA, un congénère de la famille des NAEs, interviendrait également dans la régulation du métabolisme énergétique en stimulant la sécrétion de GLP-1 et en régulant, par le fait même, la sécrétion d’insuline (Laleh et al., 2019).
Il a également été démontré qu’un dérèglement de l’endocannabinoïdome intervient généralement lorsque des perturbations prolongées ou chroniques de l’homéostasie surviennent dans le corps. Lors de telles perturbations, la production et l’action des endocannabinoïdes ne sont plus spécifiques, une suractivation des récepteurs cannabinoïdes est observée et les endocannabinoïdes se fixent à des cellules qui, à l’origine, n’étaient pas ciblées (Di Marzo, 2008). En général, dans l’obésité ainsi que dans certaines conditions telles que le diabète de type 2, une suractivation des récepteurs CB1 est observée et cette suractivation contribue au gain de poids (Li et al., 2011). En effet, plusieurs études ont démontré que les niveaux des médiateurs de l’endocannabinoïdome corrèlent positivement avec l’adiposité (Blüher et al., 2006b; Côté et al., 2007b) et des antagonistes des récepteurs CB1 ont été développés afin de traiter l’obésité et ces complications (Di Marzo, 2008).
En outre, en plus de son rôle dans le contrôle périphérique de l’homéostasie énergétique, plusieurs composantes de l’endocannabinoïdome sont également présentes au niveau du système nerveux central. Ce système de signalisation lipidique est donc étroitement impliqué, avec d’autres neuropeptides et neurotransmetteurs, dans l’homéostasie énergétique via la régulation de la prise alimentaire tel que décrit dans la prochaine section (Cristino et al., 2014).
1.3.4 L’endocannabinoïdome et la régulation de la prise alimentaire
La prise alimentaire est régulée par des processus homéostatiques et non homéostatiques. D’une part, l’alimentation homéostatique est basée sur la balance énergétique ainsi que sur la charge gastro-intestinale et vise à combler les besoins de l’organisme. D’autre part, l’alimentation non homéostatique, ou hédonique, est basée sur la consommation de nourriture pour le plaisir (Lau et al., 2017; Lutter & Nestler, 2009). Des facteurs cognitifs, émotionnels ainsi que les mécanismes de récompenses sont impliqués dans la régulation de l’alimentation hédonique (Monteleone & Maj, 2013). L’alimentation homéostatique et hédonique sont sous le contrôle de l’hypothalamus et du système mésolimbique respectivement, bien que ces systèmes communiquent entre eux afin réguler la prise alimentaire (Lau et al., 2017).
Il a été démontré que l’endocannabinoïdome est étroitement impliqué dans la régulation de la prise alimentaire en intervenant à la fois dans le contrôle de l’alimentation homéostatique et hédonique (Di Marzo et al., 2009). En effet, les récepteurs CB1 sont exprimés dans plusieurs régions du cerveau impliquées dans la prise alimentaire telles que l’hypothalamus, le système mésolimbique et le tronc cérébral (Cristino et al., 2014; Di Marzo et al., 2009; Gatta-Cherifi & Cota, 2016). Il semblerait que les endocannabinoïdes régulent la prise alimentaire en se fixant aux récepteurs CB1 afin de réguler la libération de neurotransmetteurs et de neuropeptides orexigènes et anorexigènes et renforcent la valeur hédonique des aliments (Jager & Witkamp, 2014; Lau et al., 2017; Monteleone & Maj, 2013).
Plusieurs évidences scientifiques ont démontré que le système endocannabinoïde est exprimé dans l’hypothalamus et participe à la régulation de l’alimentation homéostatique (Di Marzo et al., 2009). Chez des rongeurs rassasiés, la stimulation des récepteurs CB1 dans
l’hypothalamus par l’AEA entraine en une augmentation de la prise alimentaire (Jamshidi & Taylor, 2001) tandis que, à l’opposé, l’inhibition pharmacologique ou génétique des récepteurs CB1 induit une diminution de l’apport énergétique (Colombo et al., 1998; Di Marzo et al., 2001). De plus, il a été démontré que, à la suite d’une courte privation de nourriture, les niveaux de 2-AG augmentent dans l’hypothalamus des rats et retournent à leurs niveaux de base après consommation de nourriture, et ce, sans changement dans les régions du cerveau qui ne sont pas impliquées dans la prise alimentaire (Kirkham et al., 2002). À l’opposé, une privation de nourriture prolongée entraine une diminution des niveaux de 2-AG dans l’hypothalamus, un mécanisme adaptatif qui pourrait viser à réduire l’appétit lors de longues périodes de privation de nourriture (Di Marzo et al., 2009).
Les composantes de l’endocannabinoïdome sont également présentes dans différentes structures du système mésolimbique (c.-à-d. noyau accumbens et aire tegmentale ventrale) et ce système intervient dans la régulation de l’alimentation hédonique (Monteleone & Maj, 2013). L’activation des récepteurs CB1 et les interactions entre l’endocannabinoïdome et les voies dopaminergiques et opioïdergiques influencent le circuit de la récompense et de renforcement du système mésolimbique, ce qui augmente la préférence pour les aliments très palatables (Cristino et al., 2014). Chez l’animal, il a été démontré que l’administration d’AEA dans le noyau accumbens augmente la prise alimentaire ainsi que le nombre de réactions hédoniques positives lors de la consommation de saccharose (Mahler et al., 2007). De plus, des études ont démontré que l’exposition ou la consommation d’un aliment palatable entrainent une augmentation des niveaux plasmatiques des endocannabinoïdes chez les individus de poids normal et souffrant d’obésité (Monteleone et al., 2016; Monteleone et al., 2017; Monteleone et al., 2012; Rigamonti et al., 2015).
Par ailleurs, les médiateurs de l’endocannabinoïdome agiraient également sur des terminaisons sensorielles périphériques dans le système gastro-intestinal afin de moduler la prise alimentaire (Gómez et al., 2002). Il a été démontré que les niveaux d’AEA augmentent dans l’intestin suivant la privation de nourriture, suggérant que cet endocannabinoïde agit comme signal orexigène (Capasso & Izzo, 2008). À l’inverse, des propriétés anorexigènes ont été attribuées à l’OEA, un congénère de la famille des NAEs, étant donné que sa fixation