Secretion of a lysophospholipase D activity by adipocytes: involvement in lysophosphatidic acid synthesis.
Texte intégral
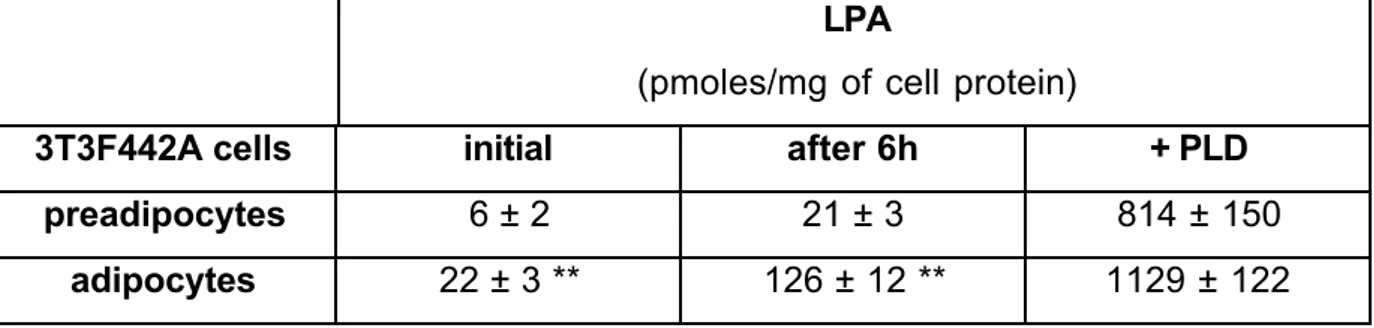
Figure
Documents relatifs
All-trans- retinoic acid represses chemokine expression in adipocytes and adipose tissue by inhibiting 1 NF-κB signaling 2 3... Abstract
(2011) New dithiolopyrrolone antibiotics induced by adding sorbic acid to the culture medium of Saccharothrix algeriensis NRRL B 24137. (2017) Efficiency of Saccharothrix
Effect of insulin, and oleic acid on triglyceride secretion by chicken hepatocytes in primary culture.. Role
Purified CD34 + cord blood hematopoietic progenitor cells were cultured in a 24-well tissue plate in RPMI1640 medium containing 10% FCS, 10% conditioned medium from MCF-7 cell
Anti- malarial activity of human group IIA secreted phospholipase A2 in relation with enzymatic hydrolysis of oxidized lipoproteins.. Infection and Immunity, American Society
(2011) New dithiolopyrrolone antibiotics induced by adding sorbic acid to the culture medium of Saccharothrix algeriensis NRRL B-24137. (2017) Efficiency of Saccharothrix
erythropoietic culture of human PB-MNCs in at minimum-essential medium (aMEM) supplemented with FBS, Epo, and conditioned medium from cultures of the 5637 bladder- carcinoma
Cathepsin-D, a key protease in breast cancer, is up-regulated in obese mouse and human adipose tissue, and controls adipogenesis.: Cathepsin D, adipogenesis and obesity...
