o '
~
.'",;'01, lH~l\IIK(lJIII$ SJ4YiIr.!.lUS:JAli J ml 5I1LI . . ,!MCGYiiiSs 41. t a 5 4 dAMa: !JIH _ i4i • ""Pi '!\t toi hi 1W4$lt rI kA . . ;&:;_, .S U N."pi,,,,--.
l
PRCDUCl'ICN, PURIFlCATlOO El' CARJlCI'ERISATICN D' HEM)LYSINES DE 'l'REPCX'ID1A HYWYSENl'ERIAE
par
BENJ!T PICARD
Thèse soumise à la faculté des étudès' graduées,
et de la rechercœ â titre d'exigence partielle
en vue de l'obtention d'un dipl✠de
Docteur en Philosophie (Ph.D.)
Départeœnt de L-ticrobiologie
campus Macdonald de l'Université M:Gill
lvbntréal, Québec 1 canada
e
B. Picardaoût 1984
- ,
..
..
....
(
Ph.D. BEIDrr PICARD Microbiologie
PRŒ>OCTICN, PURIFlCATICN El' CARACTERI~TIOO
D' HE-DLYSINES DE TREPONEMA HYŒ>YSENIERIAE
Les conditions de prcduction, en milieu liquide, d'activit€
,
hém:>lytique par des cellules proliférantes de Trepcl!'lEma hycx:1ysenteria.eet de Treponsna. imocens sant décrites.
?\
pertir d'une culture de!.'
hyodysenteriae effectuée avec l' addi ~on d'acide ribonucléique, deux hénolysines sont dérontréesdans ie surnageant. Pour chacune d' elle, une métb::xie de purification
est proFCsée penœttant l'obtention de deux prép3.ration h€rrolyt~ques
h:m:lgènes' en filtration ITOiéculaire et en électropll:?>r~se. U_ne
héIrolysine est associée à l'acide rib::mucléique (HN) tarrlis que
l'autre est associée à l'albumine sérique bovine (HP). Elles
pë!rtagent certaines propriétés cœmunes: leur synthèse est .inhil:ée ,0 ~ , par le chloramphénicol, elles ne sont ni protéolytiqùes, ni
phospholipas~ques, ni sensibles à l'oxygène; leur act\v~té ne
nécessite pas de
cat~ons
bivalents et est insensible à\Jeffet de l'EDTA; elles lysent toutes les espèces dl érythrocytes testées(boeuf, cheval, lapin, flOU ton et FOrc). Elles diffèrent par la
t
(
•
"
,
.
(('
la stabii'ité il. la t:enpl!rature et il. dilfêrents pH ainsi
t
pal; letÏPe
de sensibilité aux enzyrœs protéol}tiques. .-'~
LI isolanent et l'utilisation d' lUle souche IlU.1tante nan-hémolytique
a penni.s de démJntrer une relation entre la p:rte du pJUVOir
f
térolytique et la perte du IXJUVoir entérotaxique dans des segments
ligaturés d' intestin grêle de lapin.
, '
)\....-~~~m~"~"q."._'
•••••••.•
W.Z •• ~2.~M"M~).$KQ".i •• ~~.~M2.4!"$.! . . It ••• 2.tt.2 •• ' ... M~4Mt4.U •• aM_~MWl~"* __ KeA.'Z_4~#.;M~.~_', __ '~. ___ 4W~t._MJiI_. _ _ _ w ________ • _______ _,
<.
Ph.O. BEWIT PICARD Micrd:>iology
PlUX.L'TIOO, PURIFICATION AW CHAAACTEru:ZATION
OF HEM)LYSms FRœ 'l'REPCNF.MA HYCDySEN!'ERIAE
'!he corili. tions by which the pMuction, in liquid tœdia, of
~stances with he!rolytic activity by growin:J cells of Trepanera
hJ'C?C!Xsenteriae arrl Treponana innocens are described.
FrOn a culture broth of T. hycrlysenteriae to which ril:xj>nucleic
1
acid has been added at inxulation t.ime, t\IO harolysins are sh:lwn
in the supernatant. For each of then, a p.rrification schare allowi.n;
the obtention of an h::Irogeneous harolytic preparation by gel
filtra-tion
am
electropooresis, is proposed. One harolysin is asscx::iatedwith rilimucf acid (!lN) while the other is, associated witll. txwine
serum a1.burnirl (HP).
'l'hE:y
share sane properties: their synthesis is blockec1 by chloramphenicol, they do net have proteolytic orphos-pholipasic activities, they are insensitive ta oxygen.and the+r
activity does not require bivalent cations or is sensitive ta
,
~-.
EDTA' 5 actioni they are lytic to aH red blcx::d. ce11' s type tested
(beef, herse, pig, rabbit, sheep). They differ by'ttle size of
tpeir awarent rrolecular weight, their rrcde Qf action, the
stabi-lit Y of their activity to temperature and pH and, their pattern
of sensitivity to proteolytic enzymes.
"
,
1
1
1
f
1
1
1 11
~ 1 1 1 i , • i 1j
•
VI
'n1e isolation of a no.n-he!!olYtic nutant strain
of T. hycdysenteriae ans its use in a rabbit's ligated
ileal loop IOOdel bas s~ that there is a relation
between the loss of the hEIoolytic activityand the loss
of the enterotaxic activity.
•
,.
(
)
'.
1 t(
~ilq!1! Sb a dl
J
[/;:. ~. ~ L t" ~.f
1C
~ !.; ~r
l, t ~ '. 1 Il, _ )1. Il UUll . . . . 1 ( 1 1 %GU;;4 0 _ : un if '].
L'auteur d~sire re~ercier toutes les personnes qui
ont cQntribu~, de pr~s ou de loin, à la r~alisation de ce
travàil et particuli~remeht:
Feu le docteur Sa;ir,A. sah~, avec qui j'ai
effectué sous sa direction la majeure partie de ce projet,
1 {
pour m'avoir initié
a
la recherche en bactériolog~H desanaérobies et avoir été mon mentor.
1
Le·do~teur Réjean Beaudet, professeur au éentre 1 de recherche en baatériologie de llInstf-tut Armand Frappis+"
1
pour m'avoir guidé dans la derni~re phase de cette recherche
.
et m'avoir prodigué de judicieux conseils pour la
présenta-"
tion de cette th~se.
---Les docteurs Robert A. McLeod et Roger Knowles
qui furent, tour
A
tour, directe~r du. département de micro-fbiologie du campus Macdonald de l'Université McGill"
pour m'avo~r permis l'accês
A
une formation acaaémiquesans nulle autre pareille au Canada.
.'
..
, / 1 i •1
1
c
.
,~ ~ ~ ~ ... If~~~'i --jto:.:: ~..4-'t"1-~~'-~f-~""'f!!.t'~~""1'l!~"....j~""~"'"III.~3Ui'SIl'~:atÇCJ~~H;;;"~&.!.~fiI?,11t5~~J(j.lfll aw.
,
t .Le docteur Vincent Portelance, directeur du
..
Centre de recherche en bact~~iologie de l'Institut
Armand-Frappier, et le docteur Aur~e Beauln~s,
(lirecteur de l ' Insti tut Armand-Frappier, po'ur m'avoir
permis de participer, en tant qu'étudiant, à la vie
de cette institution de savoir et
.
d'e~seignement..
_ Madame Nicole Daigneault-Sylvestre etmonsieur Serge Durand qui furent techniciens assignés
au docteur Saheb et qui f.urent pour moi des camarades,
collègues et des enseignants très appréciés.
___ -.", 1
M~s collègues étudiants gradués du Centre de
recherche en ~actériologie (Richard 'Degré, Daniel
Dubreuil, Michel Frenette, Lucie Lafond et André Morin)
avec qui j'ai pu partager les hauts et les bas de la
vie d'un étudiant-chercheur et à qui je garde toute
-mon amitié. VIII • 1 _ _ _ _ _ _ _"
.c
"l
(
-',
A_Ids: 2 52LPiaMas . _ _ tlbU 4%1 e Il ASa a 1 1 1
!USe;
1 •
: \
"
"
.
CONTRIBUTIONS
.
A~L'AVANCEMENT DES CONNAISSANCES!
1. L'amli1iorati'on
dès
conditions de culture perme'ttant ..
la croissance et la production' h~molytique de
T. hyodysente.riae addïtionn~ d'ARN • .' '
.
,2. La mise en ~viden~e de ~'effet. de l ' addition d' ARN
"
----
---'_ ...--et d' Allli-core sur la produ~tion l1êmolytique de
.
~• l , 'il
T. ~nnocens.,
3. La mise ~ën évidence' de l'effet ,
.
de l ' addi tion' depeptone phytone. et d'ARN-core sur ·la production
h€molytique _d~ ~. hyodysenteriae.
4. L'€laboratlon d'une m~thàde de purification,de
l' hli!molysine HN.
,
.
5. La mi se en' évid';:nce de la ·p;rêsence \, d' une .
deuxi~me
,
hémolysine dahs .les su;rnagiants d~ éulture de
T. hyodysenteriae
addition~é
dJARN"
6. L'~laboration d'une méthode de purification de
Il
.
t..
l' hémolysin~ HP.
7. La mise en évidepce de la relation qui existe' entre
... ,
l'albumine sérique 'povine et le complexe' HP.
8. L'€tude comparative des caract~ristiques des hêmolysines
HN et HP. -,
9 L" isolement d'une souche mutante de -
!t.'
:hyodysenteriae" non' hémolytique. ,... o "
.
,.
'-... it
,
\
10. La mise en €vidence de la relation qui existe entre
la perte de pouvoir hémolytique et la perte du pouvoir
entérotoxique de
!.'
hyodysenteriae dans ~es segmentsligaturés d'intestin grêle de lapin.
! ,1
.
!•
x • 1;(
'.
Il
TABLE DES MATIERES
RESUME •..
ABSTRACT ...•....
REMERCIEMEN1'S .. i .
~
CON~RIBUTION
,
AU PROGRES DES CONNAISSANCES.,
, ,
.
TABLE DES MATIERES.
l
..
'LISTE 'DES TABLEAUX.
. ... .
LIS~E DES FIGURES ••
.
"..
. ...
'"... .
~REVIATIONS •••••••••• •. *8* • • • *, • • ,. • • • • • • • • • • • • • • • • • • • • • • 1. II. 'IÎido INTRODUCTION •.. ~~ •• ~ ..•. REVUE DE LITTERATpRE ••.••
.
. .
.
.
.
.
...
... 2.1 Les spiroch~tes ~G • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 2.1.1 position taxonpmique ••••• i . h2. 1. 1. 1 Le genre, Spl.roc aeta •••••.•••••••••.•••••
.
' 2 . 1 • 1. 2 Le genre '. c'tiSj!ispira • • • • • . . . .. 1 2.1.1.3 Le genre Treponema .••••.
. .
. . .
. .
.
.
. . . .
. .
.
.
2.1.1.4 Le genre Borre1i,a •••••• 2.1.1.5 Le genre Leptp~pira •••...
2.1.2 Les trépon~mes pathog~nes et non
pathog~nes ..•
. . . .
. .
.
"
, '
2.1.2.1 T. hyodysenteria~: •.
...
2.1.2.1.1 Eti~logie de la dys~nterie porcine •.•••
10 10
1.0
2.1.2.1.2 Epidé~iologie de la dysenterie porcine. 13,
(
(
XII Page 2.1.2.1.3 Aspects de la pathogénie de la dysenterie porclne... 152.2 Les toxines bact€riennes... 18
2.2.1 Types de localisations cellulaires
(Raynaud et Alouf, 1970) •••...•.••... 19
2.2.1.1 Les endotoxines de nature prot€ique... 19
2.2.1.2 Les toxines prot€iques v€ritab1es
(Raynaud et ai., 1955a) •.•••• " •• '. •••• . • • 20
2.2.1.3 Les toxines protéiques que l'on retrouve
~ la fois dans le milieu extrace11ulaire
et dans la cellule ,bactérienne pe~dant
la phase active de croissance •.••••••.•• 2.2.1.4 Le complexe toxique
protéino-" polysaccharidique de la paroi des
21
bactéries à Gram n€gatif ••••. -. • • • • • • • • . • 21
2.2.2 Classification mu1tifactorie11e... 22
2.2.2.1 Les phosphplipases hémolytiques.~... 29
2.2.2.2 Les hémolysines activées par les
r
groupements thiols.... •••••••••••••••••• 33
2.2.2.3 Les toxines non classifiées:... 35
iii l D
, ~,
• 1
r
!-' f '. ; , ~ ... ,,,"l'1ftrlfP~-n'~!~PWf;9(.lttCf''Q~~J::;;wt''-g$4i #r':;-W#)U~M.#;{~.'v."""~~lIJlt"'tt~~~WJI .. _~~~.,.:,,, ___ _ JI(
• c2.2.3 Les hémolysines chez les spiroch~tes •••••••
2.2.3.1 Le genre Leptospira ••••.••..••••••••••••• 2.2.3.2 Le genre Treponema •.••.•••••••••••••••••• III. MATERIEL ET METHODES •••••.•..••••...•.••••••••••. 3.1 Production d'activité hémolytique •••••..••••.
3.1.1 Souches ~actériennes •..•••••.••••••••••••••
3.1.2 Milieux de culture ••••••••••••..•••••••.•••
3.~.2.l Milieu solide •••••••.••.•••...•••••••••••
.
3.1.2.2 Milieu liquide pré-réduit s~ériiisé de
façon anaérobie (PRSA) ••••••• ,~ •••••••.•••
3.1.3 Conservation des souches ••••••••••••••••••• 3.1.4 Conditions de culture ••••••••••••••••••••••
3.1.5 Préparation des additifs ••••••••••••• 0 • • • • • •
3.1.5.1 ARN~ ARN-core . . . • . . . • . • . . .
3.1.5.2 Peptone phytone •••• ," ••••••••••• " " •••••••• 3.1.6 Enumération bactérienne •.•••••••••••••••••• 3.1.6.1 Compte des unités viables •••••••••••••••• 3.1.6.2 Enumération microscopique (Schenberg,
1967) . . . • . • • . . . . • • . . . • . . • . . . . • . 3.1:7 Dosage de l'activité hémolytique •••••••••.•
XIII
c "
XIV
Page
3,.1.iV • .1 Préparation de la suspension
d'érythrocytes . . . . 51
.
3.1.7.2 Détermination du titre hémolytique... 51
3.1.7.3 Calcul du tltre hémolytique... 52
'3.2 Purification des hémolysines . . • . • . . . . • . . . 5.3
3~2.l Hémolysine associée à l'ac~de
ribonucléique (HN) . . . • . . . •
a...
533.2.1.1 Centrifugation du milieu de culture •.•...
3.2.:1.'.2 Préc"ipitation acide •... , . . . • . . • . .
3; 2 . 1. 3 Précipitation au sulfate d' arrrronium . . . • . . . .
53
54 54
3.2.1. 4, ,Dialyse et lyophilisation... 54
,
3'.2.1.5 l\dsorption sur DEAE Sephacel . . . ,. . . 55
3.2.1.6 Filtration moléculaire sur Ultrogel
" il
AcA 44 . • • • • • • • • • • • • • • • • . • • • • • • • • • • • • • • • • •
3.2.2 Hémolyslne associée à une fraction
'A
56
prdtéique (HP)"... 56
3.2.2.1 Gentrifugation du milieu de culture... '56
3 . 2 . 2 . 2 préc~pltatlon au sulfate cl' arrrronium. . . . . . 57
3.2.2.3 Filtration moléculalre sur Sepharose 6B.. 5~
3.2.2.4 Stabilisat,i.on de l'act.lvité hémolytique
L
3.2.3 Estimations de la composition chimique •••••
3.2.3.1 Prot~ine ••••••••••••••.•.••••••.•••••••••
3.2.3 .. 2 Acide ribonucléique ••..• : .... , •••••••••• • ,.t
3. 2 • t3 • 3 Sucre . . . .
3.2.4 Critères d'homogénéit~ •...•••••••••••••••..
3.2.4.1 Filtration moléculaire ••.••••••••••••••••
3.2.4.~ Electrophorèse en gel de polyacrylamide •.
3.-.f.4.3 Electrophorèse en gel de
polyacrylaihide avec SOS ••••••••••••••••••
3.2.4.4 Electrophorèse en gel d' agarose ••••••• ' •••
3.2.4.5 Coloration des gels . . . )
3.2.4.6 Détermin~ion de l'identitê antigénique
xv
' g e 59 59 59 60 60 60 60 61 61 61par immunodiffusion radiale... 63
3.3 Carkctéri'sation des activités' h'êmo1ytigtres ••. 3.3.1 Cinétique de production ••••.•••••••••••••••
64 64
3.3.2 Cinétique de l'activité hémolytique... 65
3.3.3 Mode d'action des h~molysines: effet de ?
la température... 65
•
3.3.4 Stabilité des hémolysines ~ différentes
températures et, cinétique d'inactivation." 66
3.3.5 Stabilité au pH... ·67
,1
ï
-1
1 f 1 l, 1/ XVI
Page
"3.3.6 Effet àes èations bivalents et ,de l'EDTA... 68
3.3.7 Effet des phospholipides et du
cholestérol sur l'activité des hémolysines
HN et HP... 68
3.3.8 Effet de différents enzymes sur l'activité
.. h~rnolytique ••...••••..• •••••••••••• ~... 69
3.3.9 Effet du chloramphénicol sur l'évolution de
d'activité hémolytique... 71
3.3.10 Détermination du spectre hémolytique... 71
3.3.11 Estimations du poids moléculaire apparent. 72
3.3.ll.l·Filtration moléculaire... 72
3.3.11.2 Electrophor~se en gel de polyacrylamide
avec SDS •••.••••••••• • "'. • • • • • • • • • • • • • • •.• • • 73
3.3.12 Détermination de l'identité antigénique de
l'hémolysine HP par immunodif~usion
.
radiale . . . " .. ~ . . . . 74
3.3.13 Na~ure de l'activité hémolytique... 75
3.3.l3~1 Effet d~un agent réducteur... 75
3.3.13.2 Détermination du pouvoir protéolytique.. 75
3.3.1'3.3 Détermination d' une activité
phospholipasique. • • • • • . • • •.• • • • • • • • • • • • • • 76
3.4 Infection expérimentale ~ ~ se~ents
<-
ligaturés d'intestin grêle de lapin ... . 771
,
f,
~.
XVII ~ ~-"('"
.
~ -Page IV. RESULTATS •••.••. _ • . . • • . . . • • • . • . . • • . . • • • • . . • . . • • . . .,
r
4.1 Production de l'activité hémolytique ..• ~~ ••.~
79
79
f
4.1.1' T. h:l0d:lsenteriae en milieu de base . . .
4.1. 2 T. hxodxsenteriae avec addition d'ARN •...••
4.1. 3 T. hyodxsenteriae avec addition d'ARN-core.
4.1. 4 T. innocens en milieu de, base . . .
4.1. 5 T. innocens avec addition·d'ARN ••.•••..••••
4.1.6 T. innocens avec adqition d·ARN-core ...• 4.1.7 T. hyodysenteriae avec variations de la
composition en peptones du milieu de
79 81 82 82 82 96 culture . • . . . • . . . • · •....••...•.• 87
4.L.B'!~ hyodysenteriae avec variations de
l'agitation et de la température
. d'incubation . . . 4 • • • • • • • • • • • • • • ~... 91
4.l.~.1 Effet de 1'agitation dg milieu de cultu~e •. 91 4.1. 8.2 Effet de la température; d' incuba'tion ...•. ' 93 4 • l • 9 S ornrna ire. . . . . . . .. . "'.,. . . . .. .. .. . .' . . .. . . .. .. .. . . . .. . .". 9 3
4.2 Purification de l'activité hémolytique . . . • . 95
~ ~---
--~~~~--4'.2.1 L'a'ctivité hémolytique asso~iée à l'ARN (HN). ,95
4.2.1.1 Précipi~ation en milieu acide . . . ~ ..•• 96'
4.2.1.2 ?récï;:'l.tation au sulfate d'ar.r.oniun. ••...•••...•. 97
4.2.1.3 Chromatographie sur échangeur d'ions
(DEl'Œ Sephacel) ... : ... . • 1 . . 97
, ,
~)f.! l'~-;~ '''''~''.1~.'r'~l'-'l'1.~·f'i''':4..q,''-'S,t~ ~~""~M,t-,l-"""'!"'''I,~''l!~r~'''»~:J ~~".j."Iq..~~."..,.J.'i/~~~~ .. ifA~--r;~~"'''':~~~~_~-'
t.
,(
(
,
< '4.2.1.4 Filtration moléculaire sur Ultrogel
XVIII'
Page
AcA 44... . . . . ,.... 101
4.2.1.9 Homogénéité .de a préparation et
nature chimique •..••••.• ~... 101
4.2.1.6 Détermination d'un poids moléculaire
apparent. ,.. .. . ... . .. . . .. .. . . .. ... . .. .. . . . . 106
4.2.2 Indications sur la prés~nce d'une seconde
hémolysine. . . . . . . . . . . . . . . . . . 106
4.2.2.1 Cinétique d~ production des activités
ht§molytiques . . . . 109
.
4.2.2.2 Stabilisation de l'activité hémolytique.. 110
4,.2.3 Purification de l ' hémolysine Çls~ociée A une
fraction protéique (HP)... 117
4 • 2 . 3 . 1 Précipitation au sulfate d' amroniun •••••••.•••• ~ 119
4.2.3.2 Filtration moléculaire sur Sepharose 6B.. 119
4.2.3.3 Homogénéité de la préparation et
l!ature chimique ...
t. •••.••• ...
1204.2.3.4 Détermination d'un poids moléculaire
apparent . . . ~ . . . ,. . . . 124
~
4.2.4 Identité immunologique... 124
4.2.4.1 Entre HN et HP... 124
",4.2.4.2 Entre HP et le milieu de culture ••..•••• ~ 130
(-.'
,.
(
v.
•
XIX'"
,...
•
Page -'4.3.2
Cinétique de l'activité hémolytique... 1334.3.3 'Etude du mode d'action •••••••• ~.... •••••••• 140
, 4.3.4 Stabilité en fonction de
1~
température etcinétique d'inactivation... 146
.
'4.3.5 Stabilité au pH... 146
4.3.6 'Effet des cations bivalents et de l'~DTA... 150.
4.3.7 Effet du cholestérol, et des phospholipides. 151
'"
4.3.8 Effet de traitements enzymatiques •••••••••• 151
.
4 • 3 • 9 Effet du chloramphénicol •••••• ,!, • • • • • • • • • • • • • 156
4.3.10 Nature de l'activité hémolytique... 156
4.3.11 Infection expérimentale de segments iigaturés d'intestin grêle de lapin
par
!.
hyodysenteriae et!.
innocens... 164DISCUSSION ••••••••••.••••••.• ~ • . . . AN'NEXE- I . . . : 168 176 , AN'NEXE
II...
177•
AN'NEXE. I I I . . . ~ . . . , . . . l" . . . • ' . 182 ANNEXEIV...
186 1 BIBLIOGRAPHIE . . . '. • • • • • 187\
,.1
•l ,
(
,
.
~'l~ ~'~'~~f'i'" eH"""/ ~"';l<''''·(~'' , ~. ~ ... rr: .... ~~} ... :-'" ... ~,.~ rH -1 .... ~w .. "'rf!V.t'~~('!·III".f!'1~~'ti.'J...~IO,.-.. ~7't.,... ... ~~~'I"1";"~1"";.'~~,.. _" ... , t< ... ,...?"_._?' ...
-, ,
LISTE DES TABLEAUX
T~eau page
1. Classification des principales exotoxines
pro-téiques d'origine bactérienne (selon Daline et
F inch, 1 9 7 9 ) . . . 23
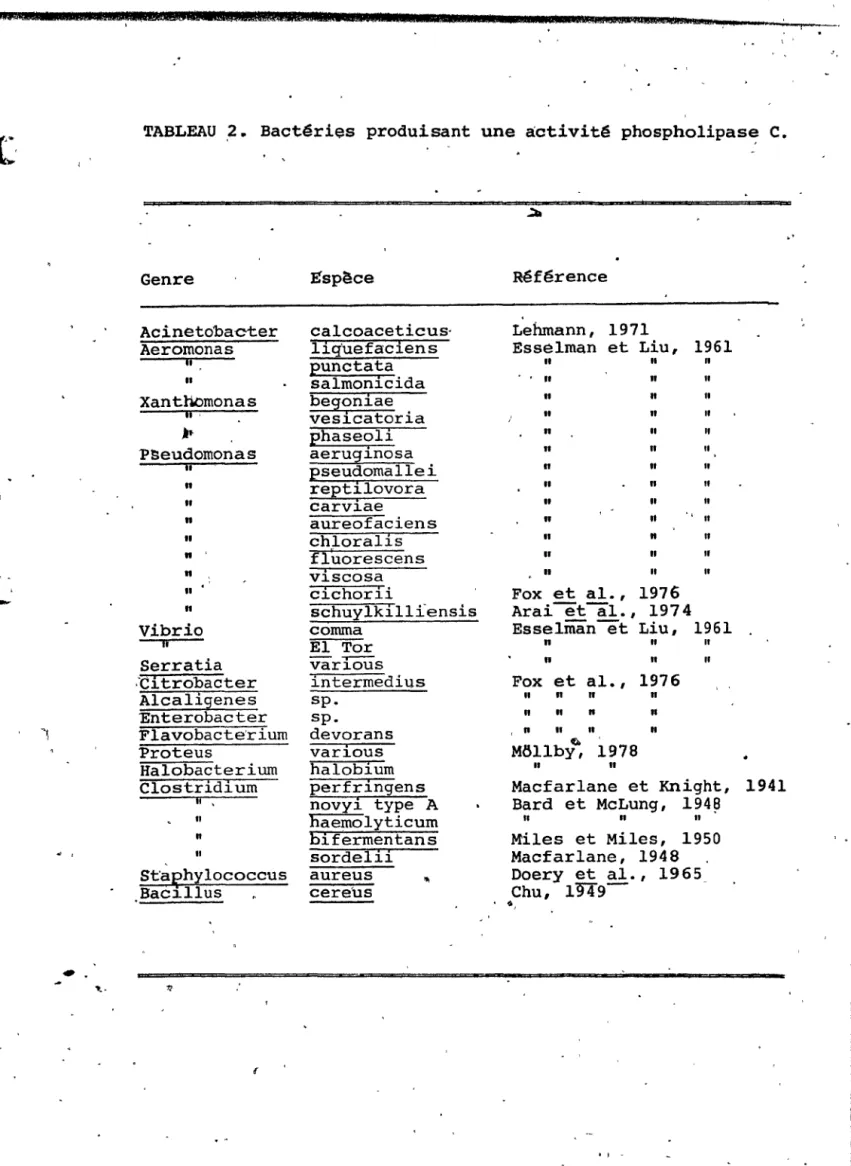
2. Bactéries produisant une activité
phospholipase C . . . 30
3. Spécificité de substrat de différentes
phospholipases C (M~llby, 1978)... 34
4. Espèces bactériennes produisant des toxines
cytolytiques activées par des groupementp
thiols (Alouf, 1977)... 36
5. Spectre hémolytique, apr~s l heure à 37°C de
.
.
surnageants de culture de différentes souches
.
'de
!.
hy-odysenteriae (A) et de!.
in'nocens (B)cultivées en milieu TSB-PRSA en présence d'ARN
(1%, plv), exprimé en pourcentage de l'acti~i
té obtenue par l'espèce d'érythrocyte la plus
sensible. . . . . . . . . . . . . . . . . . . . . . . . . . . 40
(
,-..-
...,
.
,
Tableau.
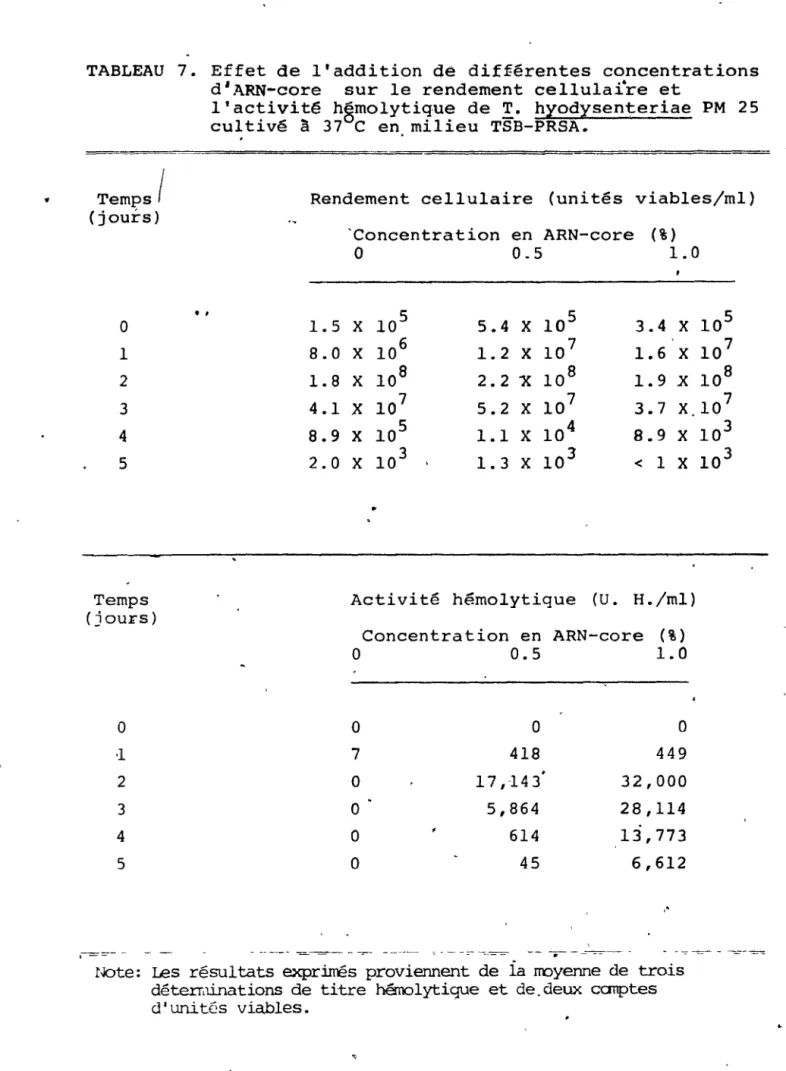
6. Effet de l'addition de différentes
concentra-7.
tions d'ARN sur le rendement , c~llula~re et
l'aètivité hémolytique de T. hyodysenteriae
PM 25 cultivé à 37°C en milieu TSB-PRSA •••••••
Effet de l'addition de différentes conc~tra~
,tions d'ARN-core sur le rendement cellulaire
et l'activité hémolytique de
!.
hyodysenteriaePM 25 cultivé à 370C en milieu TSB-PRSA •....• !
,
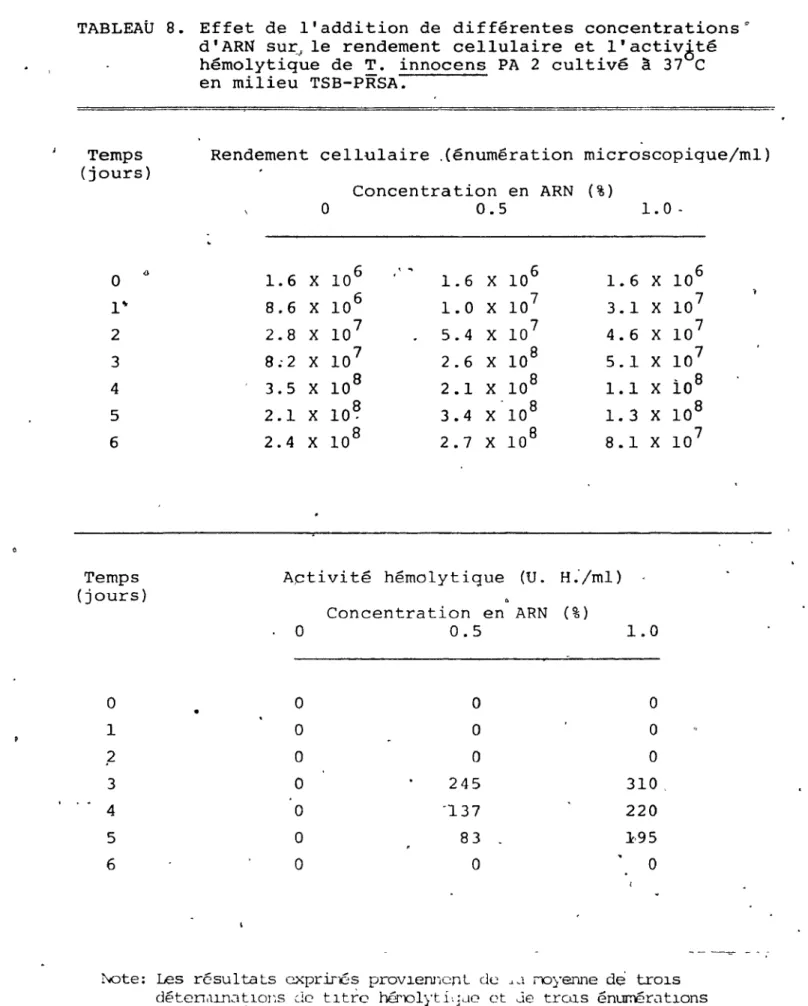
8. Effet de l'addttion de dir'férentes
concentra-tions d'ARN sur le renaement cellulaire et ",
• l'activité hémolytique de T. innocens PA 2
page,
80
"
---83
cultivé 1 à 370C en milieu TSB-PRSA •••••••••••• ~ 84'
9,. Effet de l'addition de différentes
concentra-tions dlARN-core sur le rendement cellulai~e
et l'activité hémolytique de
!.'
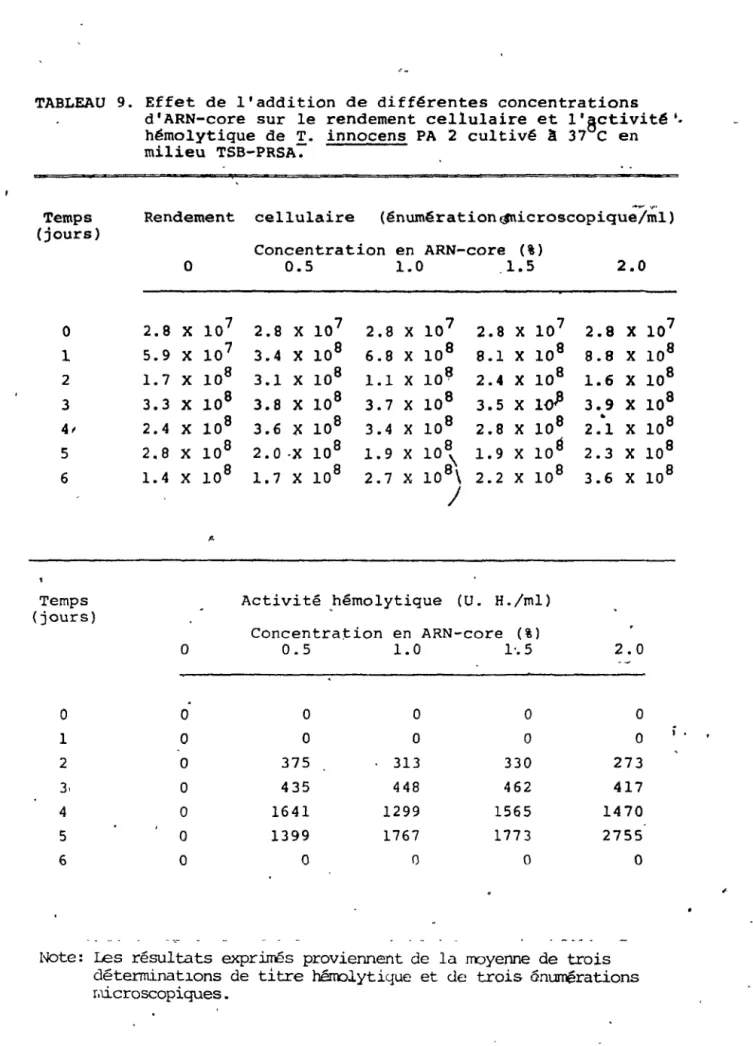
innoc'ens PA 2cultivé à 37°C en milieu TSB-PRSA... 85.
10. Effet "de la modification qualitative de la
composition en peptone (;atio phytone/
, ~
tryptone) du milieu TSB-PRSA sur le rendement
cellulaire et l'activité hêmo~ytique de
T. hyodysenteriae PM 25 ~tivé à 370C en
milieu TSB-PRSA additionné d'ARN (1%,
plv) ••••
881 l
1
J
1. ,"
.
(
Tableau
11. Effet de l'ad~ition de différentes
concéntra-tions de phytone sur le rendement cel~ulaire et
" l'activité hémolytique de T. hyodysenteriae
PM 25 cultivé à 37°C en milieu TSB-PRSA
add-i-XXII' '
page
tionné.d'ARN (1%, p/v) .••••••.••.•••...•• ~.... 90
12. Effet dé l'agitation sur le rendement cellulaire
.
et l'activité pémolytique d~
!.
hyodysenteriaePM 25 cultivé à 37°C en milieu TSB-PRSA
additionné d'ARN (1%, plv) ••... , •••.• , ..•.•• -, ,92
13. Effet de deux températures d' incubation
.
su~ le'-- . .~rendement cellulaire et l'activité hémolytique
de T. hyodysenteriae PM 25~ult~vé en milieu
TSB-PRSA additioh.né d' ,
ARN
(1%, plv) ••••••• '. ~ • • 94~
14. Purification de l'hémolysine HN de
---
!.
hyodysenteriae PM 25 •••••••••••••••••••••••..
15. Effet'de la gélatine et du glycérol sur la
stabilisation de l'activité hémolytique
..
HP de9,8
"
.
!.
hyodysenteriae "PM 2 5 . . . 11516. Effet comparatif de différents sels sur la
,
stabilisation de l'ac~ivité hémolytique de
T. hyodysenteriae PM 25 .••••••••.•.••••••••••• ' 116
--.,...
'. o •.
' ,.
.
--, , 1 • _ 0t
" 0',(
..
", ,.
~ -~~-~----.... -~-~--~- --~-~._-~-.
__
... _---~-- -..-.- -
.... --~-- -~ ~---.-.-...---XXIII Tableau page , 17. Purification de l'h€molysine HP de 18,
T. hyodysent.er i'ae P~f 25~' ;-. : ••••••••••• ~ ••••••• oSpectr~ h€molytique des hémolysines purifiéès
HN et HP deoT. hyodysenteriae PM 25, exprim€
en pourcentage de
l'activi~)hémOlyti~ue
obte-nue par l'esp~ce d'érythropyte,la plus $ensi~
ble apr~s incubation d,une heure à 37oC •••••••
19. Etude du mode d'action de l"hémolysine HN de
T. hyodysente'riae PM 29 à différentes
tempé-ratures en fonction du .temps .•••••••• ' ••••••••••
20. Etude du mode d'action de l'hémolysine HP de
T. hyodysenteriae PM 25 à différentes
tempé-118
132.
142
ratures en fonction du temps •• c • • • • • • • • • • • • ~ • • • "143
21. Etude du mode d'action des hémolysines HN et
HP,de T. hyodysenteriae PM 25 à différentes
températures en fonction du temps... •.••••••••• 144
22. Evolution de l'activité hémolytique des
hémo-•
,
lysines HN et HP de'!. hyodysenteriae PM 25 ___ , .
"- .
en fonction de la tem~érature ••••••••••••••••• 147
23.- Effet des cations bivalents et de l'EDTA sur
l'activité hémolytique des hémoly~ines RN et
HP de, T. hyodysenteriae PM 25 •••••••••.• ~ •••• '. 152
..
l' Il 1 1 1,1
1·,
./
l
(.
{)"
XXIV Tableau pag~24. Effet du cholestérol et des phospholipides sur
l'activité des hémolysines HN et HP de
T. hyodysenteriae PM 2 5 . . . 154
25. Effet de traitements enzymatiques sur l'activité
des hémolysines HN et HP de
!.
hyodysenteriae~-PM 25 . . . "._... 155
26. Effet de la cystéine sur l'activité des
hémo-lysines RN et HP de
!.
hyodysenteriae PM 25 •.. 16327. Infection expérimentale de segments ligaturés
o d'intestin grêle de lapin p~r T. hyodysenteriae
et T. l..nnocens ... .
.
165 ~I1 \ .,
t.
Figure 1. 2. 3. 4 •.LISTE DES ,FIGURES
...
Profil d'élution de l'hêmolysine HN de
!.
hyodysenteriae PM 25 sur DEAE Sephacel •••••Profil d'éluti~n de l'hémolysine RN de
!.
hyodysenteriae PM 25 sur Ultrogel AcA 44 •••..
Electrophor~se en gel de polyacry1amide (10%),
....
pH 8.9 de l'hémolysine HN de
!.
hyodys~nteriaePM 2,5 (50 llg de protéine): apr~s fil'Cration
moléculaire sur ,Ul trogel AcA 44 •••••••••..••••
t
Electrophor~se en gel d'agarose (1%, plv) A
pH 8:4 de l'hémolysine HN de T. hyodysenteriae
PM 25 (50 llg de protéine) apr~s filtration •
)
moléculaire sur U1trogel AcA 44 ••• , ••••••••• ~ ••
page 100 103
.
, b • f ' ,105 .,5. Détermimitïon du poids "moléculaire él;pparent de,·,.J' '"
, ' ,
, , " f
1 • hémolysine HN de T. hyodys'ent'eri'ae PM: ,~5 pàr
~iltration ~ moléculaire sur Ultrogel AcA 44 ••••
j \ t 108
6. CinétJqu~ de production des hêmolysines HN et • , p
.
I(P--<le
!.
hyodysenteriae PM 25 cultivé '~ 37°Cl ,
sans agitation en mili,eu TSB-PRSA additionné
d'ARN (1%, p/v) •••••••• ' ••••• ~... 112 ; , , o '. , (
î
l,
fi '
1
J
1 1i
1 ! 1i '
(
(
,
" • ' " ,. Figure7. Electrophorèse en gel d'agarose (1%, pH 8.4)
.
,de la production hémolytique représentative
d'une cul t'ure de!. hyodysenteriae PM 25 dan,s
un milieu TSB-PRSA à 370C sans agitation,
XXVI
page
sans (A) et avec addition d'ARN
(B)...
113B. Profil d'é1ution de l'hémolysine HP de
\
!.
hyodysenteriae PM 25 sur Sepharose 6B... 1229. E1ectrophorèse en gel de po1yacry1amide (10%,
plv) avec SDS de L'hémolysine HP de
~
!.
hyodysente~iae PM 25 (50 ~g de protéine)après fi1tratrop moléculaire sur Sepharose 6B. 123
10. Détermination- du poids moléculaire apparent
de l'h~mo1ysine'~p de
!.
hyodysenteriae PM 25par filtration molé~u1aire sur Sepharose 6B •..
Il. Détermination dû poids moléculaire apparent
de l'hémo1ysi~e HP de !. ?yodysenteriae PM 25 par électrophorèse en gel de po1yaery1amide
126 .
'avec SDS . . . t • • • • • • • • ~. • • • • • • • • • • • . . . • • • • • • • • 128
1 ~. 1
12. tcienti té i~uno1ogique des hémo1y:sines HN et
, HP de T. hyodyqente+iae PM 2,5 .•• -;- ~ • '" ••• '.. •• • • . 130
.
, \ ".
,,J'\W·'!':1~~,""~~m:t, • .tW~~J,«W\iellJ!J!II!II".j(illlt~J~($1II""~1"\>i'1Jl11l!111!11!f1~~"':'I'~'"~~_" .... ,,; " ... _.~" __ .~'"'_
Figure
13.
"
!dentité immunologique de la fraction
anti-génique de l' hémolysine HP de
!.
hyodysen'teri'ae'~' XXVII
" pag~
'PM 25 •••...•..•••...•.•.•...••.••.... ft • • • • • • 131
14. Cinétique de lyse d'érythrocytes de iapin en "
fonction du temps par l' hémoly~ine HN dd'e
!.
hyodysenteriae PM 2!i à 37°C •.••••••.• (. •• •• ' ',1'3'515. Cinétique de lyse d'éry~hrocytes ?e lapih en,
fonction du temps par 1 'hé,molysine HP de _
!.
'hyodysenteriae,PM 25 à 37 C . . . .. o16. Stabilité à différents pH des 'hémolysines HN
1
et HP de T. hyodysenteriae PM 25 apr~s 15
,
minutes ~ 25°C ... • ' • . ~ ... .
17. Effet ~u chloramphénicol sur l'évolution de
18.
. l'activité hémolytique de T. ,hyodYsenteriae
" " , d
, PM 25. cultivé en milieu TSB-PRSA ~ ,37 C.~ •••••
Effet, du chloramphénicol sur l'évolution de
l'activité' hémolyti~ue de T. hyodysenteriae
PM 25 cu~tivé en milieu TSB:PRSA additi~é
o d'ARN ,-(1%, plv) A 37 'c ....•••••••....•••••••••
J
138 , 158 160 .' ,l
" .. ifl." ' . '! Jf'., ~";~ "'tif '. ,
(
• b Figure19.' Cin~tique d'inactivation il 37°C de l'activité
hémolytique de sprnageants de culture de
!.
hyodysenteriae PM ~5 cultivé en milieuTSB-PRSA et en milieu TSB-PRSA additionné
d'ARN (1%, plv) . . . .
..
, ' XXVIII page 162., (
.'
, .1 t -~ ---~- -~ .. --_---.... ,...- -.' ABREVIATIONSARN: acide ribonUCléiqur ~
ARN-core: fraction non aigestible par la·~ibohücl~ase
pancréatique de l'acide ribonucléique
DEAE: diéthylaminoéthylcellulose :t.
EDTA: acide (éthylènedinitrilo)-tétraacétique
PBS: phosphate bufferea saline/tampon phosphate salin
PBSA: PBS additionné d'albumine
PRSA: pré-réduit stérilisé de façon anaérobie
TEMED: N,N,N'
,N'-tetr~thYlétbYl~ne~ne
Tris: base Trizma (Sigma)'
TSB: trypticase soy broth
.
.
.SOS: dodécy~ sulfate de sodium
J • ,"
.
"..
, , , ~ ,.
,.
":-.
", .' :.
'.
.
-.'
1 1 • •. 1 ,.
,..
'.
" .'..
" ?
.
(
..
., I. INTRODUCTIONLes tréponèmes forment un groupe de bactéries anaé-robies dont la majorité des espèces sont des const}tuants normaux de la flore buccale et intestinale de la plupart des espèges animales, y compris l'homme.
Au niveau du gros intestin, la présence de spirochètes a été décrite chez l'homme (Harland et Lee, 1967), le singe
(Takeuchi ~ al., 1971), le porc (Taylor et Blakemore, 1971),
le chien (Leach et al." 1973), le rat (Davis et al., 1972)
.et la sO'..1ris (Savage et al., 1971). La localisation dé ces
bactéries dans l~intestin varie selon l'espèce an~male= chez
la souris les spirochètes ne sont"pas associés aux ce,ll ules
épi théli-ales (Savage et al., 1971; Davis et al., 1973) alors
•
que chez le singe et le rat l'association est superficielle
.
';. (Takeuchi' et al., 1971; Davis et al.,~ 1972) et. que chez le
porc, ces gçrmes sont pré~ents à la ~urface et dans les
cellules épithéliales (Taylor et B1akemore, 1971; GlOCr
et ~., 1974).
1.
De tous les tréponèmes in~estinaux CORnus, un seul,
Tr~ponema hyodysenteriae est ~c~nsidsré pathogène CSmibert,
..
1971; Harris, Glock, Christensen et Kinyon, 1972; Harris,
.
"..
Î
1 1 1 i ,f
1:
~,,; '{ t-e
" , ~~l
i(
, ,1
Kinyon, Mullin et Glock, 1972; Smibert et Claterbaugh Jr.,
1972; Harris# 1974; Kinyon et Harris, 1974). En effet, les
.'signes cliniques et les l€sions caract€ristiques de la
dy-senterie porci~e aiguê ont pu être reproduits par
inocula-tion orale de cette bactérie 'à des porcs conventionnels et
.
'''''',
qnotobiotes (Taylor et, Alexander, 1971; Glock et ~arris,
1972;' Harris, Glock, Christensen et Kinyon, 1972; Akkermans ,
et ~ornper, 1973; Hamdy et Glenn, 1974~ Hughes ,et al., 1975)
et, on a retrouvé ce germe en grandes quantit€s au niveau dû
\ côlon et du caecum des porcs malades. Les travaux r€cents
tendent toutefois â démontrer que cette pathog€nie peut @tre le réspltat d'un synergisme entre!. hyodysenteriae et
d'autres composantes de la microflore intestinale (Meyer
et al., 1975; Brandenburg et al., 1977; Harris et al.~ 1978;
Meyer, 1978; Whipp et al., 1979). Nonobstant les progr~s
réalisés dans l'élucidation de l'€tiologie, peu de faits
nous permettent d~ comprendre par quel(s) mécanisme(s) cette
maladie est induite. Les différents mécanismes qui furent
sugg€rés pour expliquer l'initiation de cette colite sont:
l'invasion des cellules épithé~ia.les par le spiroch~te, la
production de cytotoxine par le trépon~me ou d'autres
bacté-ries anaérobies et, finalement l'initiatio~ de
l'infiltra-tion de la muqueuse par une toxine provenant du trépon~me
ou d'une autre bactérie (Taylor et, Blakemore, 1971; Glock
l '
~t HarFis, 1972; Kent et Moon, 1973; Hughes et al., 1975;
wilcock et Ola der, 1979).
L'hypothè e considérant la présence d'une cytotoxine
e'st basée·' sur 1- apparition souda:ine de nécrose coagulative 3
de la muqueuse, restreinte aux zones où i l existe une
pro-lifération massive de spiroch~tes et, auquel s'ajoute l~nca
paciœ de démontrer l'invasion de cellules saine~ par les
spiroch~tes ou d'autres bactéries. Il a été Suggéré que la
nécrose es~ restreinte à la partie superficielle de
l'épi-thélium parce que des variation subtiles du
microenviron-nement côlonïque permettait la présence de
!.
hyodysenteriaeet de sa (ses) toxine(s) à des concentrations adéquates,
seulement dans les zones 9 environnant l'épithéliun
superfi-ciel (Kent et Moon, 1973; Hughes et al. i. 1975)·.
L'hypo-th~se d'une invasion cellulaire est basée sur la
démonstra-tion de la présence de spirochètes à l'intérieur de
cellu-les épithéliacellu-les du côlqn en dégénérescence (Taylor et
Blakemore, 1971; Glock et Harris, 1972). Enfin l'hypoth~se
émise concernant l'initiation de l'infiltration de la muqueuse
causée par une (des) .toxine (s) refOse sur l'observation fréquent~' de caillots dans les vaisseaux sanguins de la ml..XJUè~e côlonique· de fOres atteints de dysentet<ie fOrcine (Kent et M:lon, 1973).
,
~;
.
t
~ , ~.
J(
1
mm.II,."
IlU
:W'SJliiU4Mm 2i2!tlt5ltBiI •• . .~ S"'tii!iSMIL 1 JIBI " • ---M!II _ ,aP!J~$.I. _ _ H!U#J . . :a4i4?U"""h&Zj;i
.-/
4
De plus, considêrant le fait qUi
!.
hyodysenteriaeest hêmolytique en cu~ture sur g~lose au sang et que
plusieurs auteurs ont êtabli une corr~lation entre le type
hémolytique de
!.
hXQdysenteriae et son pouvoirentéro-pathogène (Taylor et Blakemore, 1971; Meyer et al., 1974a; Meyer et al., 1974b; Meyer et al., 1975; Kinyon et al., 1977), il semblait pertinent d'étudier le pouvoir hémolytique de
T. hyodysenteriae pour éventuellement en caractêriser le mode d'action et préciser son implication au niveau de la
pathogénicité associée
A
l~ dysenterie porcine.Le cheminement logique dans l'~tude d'une tox1:ne
bac-.
têrienne peut être subdivisê en diff~rents aspects (Arbuthnott,
1978) :
• ' >
(1) la production in vitro de grandes quantités de toxine;
(2) la purification et la caractêrisation de ~ toxine;
(3) le développement d'un bioessai satisfaisant;
(4) l'élucidation du mode d'action au niveau 'physio-j
1
logique, cellulaire et mol~culaire~
(5) l'évaluation du rôle de la toxine dans la pathogênie.
•
..- ,. i"""~'r·'t .. WIJ.."""",A"it,.;",:~rr~~~'~~ '~·:,~~>' ... j-:r""}.}\~",~t-t.f"~~.lJ'~~~""~~~~",.~~~~~"~'~~~!'il.,.~H~~~~'W/Jf'I~'!\II,,*~d'M#iW""""Q""''''Mi''''. ___ I>I_' L _ _ :"',
1 .
)
/
Dans ce travail, notre attention a surtout porté sur la production, la purification et la paractérisation de
l'activité hémolytique associée à T. hyodysenteriae,
éta-pes essentielles et préliminaires aux études concernant
5
le mode d'action êt l'implication possible de la toxine dans la pathogénie.
" 1
•
"
II. REVUE DE LITTERATURE
2.1 Les spirochètes
2.1.1 Position taxonomique
C'est )par leurs ~aractéris'tiques morphologiques que
les spirochètes forment un groupe bactérien bien distinct.
On les d~finit comme des bactéries élancées, flexibles et
pouvant avoir une forme hé1ico!da~e ou ondulée. Ce sont des
organismes unicellulaires dont la longeur se situe entre
5 et 55 ~m et dont le diamètre est compri~ entre 0.1 et
3 llm.
L'u1trastructure telle que révélée par la microscopie
électronique a pu mettre. en évidence que to~s les spirochètes
,.
ont en commun les catactérist~ques morphologiques suivantes:
la cellule expose au milieu·externe une membrane multicouches
I~
que l'on désigne membrane ou enveloppe externe, le peptido-glycan et la membrane cytoplasmique sont localisés sous
l'en-veloppe externe et sont ~ignés par le tenne général de cylindre
protoplasmique. C'est entre l'enveloppe externe et la couche'
de peptidoglycan que l'on retrouve l~ caractéristique unique
\
1
(
"
,
..
, '
7
des spirochètes: la localisation de leurs flagelles. Ces
flagelles sont des organes internes, situés entre le cylindre , , . ,
, ,
protoplasmique et la membrane externe. Tous les spiroch~tes
sont pourvus d'au moins' un flagelle or~ginant en ~osition
,
.'
subterminale de chaque extrémité du cy+indre' protoplasmique
qui se rejoignent et s'entrecroisent
térie (exception faite des leptospires dont les flage~les
se rejoignent au centre mais ne s'entrecI:Oisent pas)
(Johnson, 1977).
, ,
"
, ,
Les spirochètes sont un groupe tr~s h~tér6g~ne au n~
veau de l' habitat, de la structure et de la phy~iologie. Les
.
spirochètes sont préserltement classifi~s dans ~n ordre,
" 'J
Spirochaetales -ne comprenant qu'une fam,i,lle, Spiro6haetaceae.
1 •
. ' • t •
Dans cette famille, on y regLüupe les cinq genr~s, suivants:
Spirochaeta, 'Trep:mema, <:ristis;eira, 'Bdrrelia et ,:~tospira. "
La définition des genrés fut effectuée sans appliquer les . mêmes caractèrés conune critère de différentiation car, la
JI
très grande diversité existant
aJ
niveau de la ,morphologie,
" •
et de l'habitat était amplement suffisante pour éviter toute confusion. ( \
:'.
" ' " t • , 'f , ' \f
,
" t î 1 { ! 1 _ f I&~."(.
".
... ".~~~",-
~&!
1 '°0 8 ,.
~ -'!l ;.r
\...
2.1.1.1 Le genre SEirochaeta-Le genre Spirochaeta est formé de bact€ries de Ipngeur
compr i se en tre ~ et 500 um par 0.2, à O. 75 PJll de .diam~tre; ce
sont' des bactéries anaérobies facul ta'tives que l'on peut
re-trouver à. l'etat nattlrel dans les eaux salées et douces
con-~ t " f7 , ..:
tenant du su/fulre d'
hydrog~ne'
(habituellement\, d..ans les €gouts b _ou les eaux polluées).
2.1.1.2 Le genre c;ristispira
..
Les bacUiries formant le genre .Cg:stispira sont, de
longeur comprise e'ntre 30 et 150 llm.par O.~
à.3
lJ~,dedia-,',
mË!tre. Elles ont de 3 il 10 circ:onvolut;ions comp1~tes. Dans
o
.
-,.
.
, les sÎécimens vivants, des inclusioI)s ovoldes et des touffes
de flagelles peuve~t êtJ:"e observées par microséopie en
con- .-traste de phase. Ce s,ont des commensaux que l'on retrouve
, 1. ...
habituellement associés à des mollusques.
, 2. 1. 1. 3 Le genre Treponema
:~s
'memb\s du genre Treeonema ont des longeyu:s1.
. , . > , ' "comprises entr~ 5 et 15 llm et un diam~tre variant entre
0.09 il 0.5 pm. Ils sont d~pch.p;vus de catalase et d'oxidàse,
1
.
ana€robies, commensaux ou pq.rasites, certains JSont pathogènes.
\ il
(
---(
2.1.1.4 Le
genre~Orrelia
' - A l'int~r~eur du 'genre Borrelia nous retrouvons des
bactéries de longe urs comprises ent~ 3 et 15 ~m pour un
diamètre de 0.2 à 0.5 pm. Ce sant des bactéries anaérobies
et parasites. Il existe des espèces p~thogènes et le mode
de transmission s'opère généralement par des tiqqes et des
.,
poux.
2.1.1.5 Le genre Leptospira
Les bactéries du genre Leptospira sont de longeur
comprise entre 6 7t 20 ~m et ayant un diamètré d'environ
0.1 pm, ce sont des bactéries· spiralées ayant une .longeur
9
d'onde relativement courte par rapport aux autres spirochètes.
, I~s peuvent avoir des extrémités courbées ou en forme de
cro-cb§t. Ce sont les seuls spirochètes aérobies stricts. La classification des espèces est effectuée par sérotypie
•
•seulement et, on retrouve certains sérotypes à l'état libre,
-d'autres paras~tes et certains sont pathogènes.
Les éléments de classification que~nous avons énumérés
proviennent du,IIBergey's Manual pf Determinative Bacteriology",
, _ _ _ ~~ _ _ _ _ ~ _ _ _ _ ~_J'M.BSu_; _ _ b ___ '_O~'W"· __ Jl_Id~b_J _ _ _ _ L_. ____________ ~, _._. ________ __ , J ---:;----: : 1
(
(, , ' , , " '..
, ', Le genre Treponema c,omprend 13 espèces plus' ou moins
bien d,éfinies~ Parmi des. ,espèces T~ p'a'llid\,1In,
!.
,.pertenue,1', , , ' ~ . . ,
-!.
carateum et!.
paraluis~cuniculi sont 'pathogènes etnon cùltivables tandis que
!.
hyodyse~teriae est pathog~ne ,et cultivàB1e. Les autres. espèces sont cultivable~ et
con-sidérées· parasites ou pathogène$ d'association, ce sont:
T. phagedenis, T. macrodentium,
T:
refringens,!.
dent,icola,T. orale, T. sC01iodontum, T. vincentii et T. innocens.
2.1.2.1 T. hyodysenteriae
6
2.1.2.1.1 Etioiogie de la dysenterie porcine
La dysenterie porcine a été décrite'pour la premi~re
fois dans -la littérature par ~hiting, Doyle et Spray en 1921.
o ,
Pendant près de 50 ans , les chercheurs ,furent da,ns
l'impos-sibilité d~identifier correctemènt l'agent étiologique
res-'ponsable de cett€ maladie. Puis, vers la fin de 1969
com-,...
, menca la publication d'une série de,'notès exprimant la
~
----
'conviction des auteu~s au sujet de l'association probable
de spirochètes dan,s l ' étiologie ~e la dysenterie porcine
(Vall~j:o, 1~69.; Blakemore et ~aylor, :).970; Espinase et
1
.1
l
.
,, ,~ ,
.. ,~ '":;;"~.f/"'~1-'1.~r!I( ~~ . . . ;' -:-< ,~l,."o;-j\.~ '! .. ~r.,\~-~.~fI,'J!,' <-~~~\ "~/"ôi<~":~~P~l't'~Y:"~~""'~~~ ~tff~"t'i<~~"'\~"",~~~ ... ~ll!\~'!:'('mn.~,,,,_ ... ; ___ 'ffl~ _ _ Jo.,. _ _ ... _;~ ____ _
Redon,
,
1970; Roberts et Sinunon, 197 0 . l 1 ~ ~" Taylor, 1970; Todd11
et ~., 1'970). Da!ls certa~ns ,cas, ils furent il même de
culti-,
ver des spiroch~~es proyenant de mati~réS féca~es de porcs
, ,
mal(eS (Ta~lor, 1970;, Todd et
j
al., 1970) mais i l ' faJ,lut·
attendre la publication en 1971 par Taylor et A~exa:t:lder
d'un article" re"latant la ,r~production" des symptômes et des
lésions de la dysertterie"porcine chez les porcs par l ' ad-,
•
ministrat.ion per' os d: une cu1;ture pur~'
Cl·
un spirochète. Cette,expérience fut reproduite maiptes fois chez des porcs normaux
• ,
.
et ,g'notobi<;>tes (Hatris, G10ck, Christen~en et., K~nyon, 1972;
, ,
Akkermans et Pomper, 1973; Brandenburg et ~. '1977;, Hamdy e1;:
__ . . . a
-Glenn, 1974;, Hughes' ~ al,., 1975). Les c(;:mnaissances'
progres-sant, il' apparu c1aiiement que même si la'
pré,senc~
d' 'àutres 'bac~éries était nécessaire pour permettrè a~ spiroch~tè d~
causer l rinfection, l~ seul micororganisme essentiel pour lq
,
transmission de la dysenterie porcine d'un porc il ,~' autre "
était le spiroch~te (H,arris, Glock, Christense~ et Kinyon,
1972; Branden~urg 'et al., 1977; Meyer et aL,. 1975)'.
Ce spiroch~te,fut nonuné Treponema hyodysenteriae
, \
(Harris, Glock, Christensen et Kin~on,
" ; ;' j ~ i ' f .~ , " , tl ~1I8<lJ!m',4i_ni&illJR!lt4 .. H"*",,,l§! .1I1J1j!.il!'i.iiiV".:;a4H!~!
,!
a ~ . , 1 • , . \ 4;~êçf_.mt; as.; "". ;:'\t4Yl$J;_4MIt;t#(~""'.'1""""'f~"_III111"" _ _ • _ _ _ .. __(
s1
'traditionnellement on parlait de tr~pon~mesnon-pathogènes et cul~ivables et de tr~ponêmes patpogênes
et non,-cultivables, avec
!.'
hyodysenteriae, ce fut lepremièr exemple d'un tr~pon~me pathogêne que l'on pouvait
cultiver in vitro (Taylor et Alexander, 1971>". Parmi les
,',
caractéristiques culturales de ce ge~~e ana~robie, les plus
remarquables sont celles de former rarement des colonies
~ ia surface d'un milieu gélosé et de produire une
hémo-lyse clai~e de type bêta
Ca)
(Harris, Glock, Christensenet Kinyon, 1972).
12
•
Il a été d~montré que l'on pouvait isoler des
±r~pon~-\
mes morphologiquèment similaires
èt
, antigéniquement àpparentés.
.
,
à,!.',hyodysenteriae (Harris et Kinyon,' 1974; Hudson et-a1.,
1976; Rinyonet al. 1976). Ces tr~ponèmes .différaient de
!,.
'hyodysenteriae parce q~' ils n'étaient pas pathogènes(Taylor et Alexander, 1971; Hudson et ~. 1976; Kinyon
~ 'a1. ,. 1977) et qu' ils ~taient', dotés d'une plus faible
.
'activité hémoiytique sur gélose au sang (Taylor et Alexander,
1971). A présent, il est 'génér'alement reconnu qu' il exi~te
chez ces tréponèmes une corrélation directei entre le pouvoir
• 1
entéropathogène et le type d'activi~é ~é~olytique produite'
sur une g~lose au sang (Rinyon et/.al., 1977; Kinyon et Harris,
1979).
.
1 ' 1 " ' ,?~~""'-_·f~~~~.~"~J/\ .. ~~:l_!t~""f1I~!f""'l""~~:~';$~~·~I\~'1~i'_"'~,.,t~.'.*~i'&k Lb AII'J~~t""'l;S;M't"JIl)""."'fII"' _ _ _ _ _ _ '.c
.
.
(
13 ~Les tr~ponèmes non-pathogènes peuvent se retrou~er
chez la grande majorité des'porcs (Hudson et al., 1976;
-
,Kinyon ~ al., 1976~ Joens et al., 1979). Il a été proposé
que, ce tréponème non-pathogène sott appelé Trepon'ema i'nnocens
(Kinyon et Harris, 1979). La nomination du trépon~me
hémo-.'
lytique non-pathogène au rang d'espèce distincte est basée ,...
sur trois critères principaux: l'absence d'entéropathogéni-' cité (Taylor et Alexander, 1971; Kinyon et al., 1977;'
Kinyon et Har.ris', 1979); l'activité hémolytique plus faible sur gélose au sang (Taylor et Alexander, ,1971; Kinyon et al., 1977; Kinyon et Harris, 1979) et, le peu d'homologie de son
• _' t
acide désoxyribonucléique avec les autres trépon~mes (Miao
~ al., 1978; Kinyon et Jiarris" 1979).
f
1
,2~1.2.l.2 Epidémiologie de la dysenterie porcine
La qYsenterie.porcine affecte surtout les porcs
pesant entre 15 et 70 kg, quo~que l'on puisse rencontrer
.
la .maladie' chez des' porcs ~ l' allaitem~nt et les adultes
(Harris et Glock', 1981).
La malad,ie est transmise principalement lors, de l'in-gestion par des porcs sains, de matières fécales provenant de
1
}
.
~ - j : i -• , î· , ~ 1; ~ ~.J
~ ~f·
~ l~OIA Il 1 2 bru $1111111 . . . l b t 1
la
Cd li bd J :_UItllUIBU211 'Elut dl &!llMBllla ia WSi.4h*(
(
.~
'1
porcs malades ou de porcs sains porteurs de!. hyodysen~~riae
(Songer et Harris, 1978). La morbidit~ peut atteindre 90%
et-la mortalité 30% selon le succ~s du traitement. Dans 'un
troupeau non trait~, la mortafité peut atteindre 50%
(Harris etG10ck, 1981).
La persistance de T. hyodysenteriae dans les mati~r~s
fécales de porc diluées au dixi~me dans de l'eau a été
éva-luée à 61 jours si elles sont maintenues à' SoC et à 7 jours
~ 250C ,(Chia et Taylor, 1978). Dans l~ sol, le germe a pu
conserver sa viabilité pendant 18 jours à 4°C (Harris et ,
G10ck, 1981).
'"
Le porc n'est pas le 'seul hôte possible pour
T. hyody~enteriae. Glock, Kinyon et Harris (1978), ont
dé-montré qu,'après gavage,. on peut retrouver
!.
hyodysenteriae.~ _______ 1'
-chez le chi~endant 13 jours et chez l'oiseau pendant
8 jours. Une mouche peut transporter le germe viable pendant
•
4 heures (Harris et G10ck, 1981). Chez les rongeurs, le rat
peut conserver le germe pendant 2 jours, tandis que la ~ouris
peut porter cette période à 100 jours (Harris et Glock, 1981) •
(
"",. 15
,\
2.1.2.1.3 Aspects de la pathogénie de la dysenterie porcine
Même s'il est communément accepté que la dyse~tèrie
. porcine résulte, de la p~t d~ la proli.~ération de
!.
hyodysenteriae dans ~ tractus intestinal de porcssuscep-tibles, p'eu de données nous permettent une comprél1'ension
',[
du mé~anisme par lequel la maladie est induite.
Les transformations de l'ultrastructure intestinale 'pendant les premiêres manifestations de la dysenterie porcine
ont été caractérisée (Harris et Glock, 1981). Au début, on
observe l'apparition'd'une grande quantité de spirochêtes ~
la surface de la lumiêre intestinale du côlon et dans les cryptes. Par la suite, les cellules épithéliales adjacentes présentent des lésions incluant la destruction des microvilli,
le gonflement des mitochondries et du r~ticulum endoplasmique
. /
ai·nsi qu'une densité accrue de ces organelles. Au fur et ~
mesure que les dommages .prennent de l'importance, les cel-Iules épithéliales s.e rapetissent et deviennent sombres.
!.
hyodysenteriae' envahit les cellules épithéliales, les.
"cellules en gobelet et la lamina propria. Les tréponêmes
peuvent être observés en grans nombre (quelquefois en ,
g+appes) à l'intérieur de certaines cell,ules épithéliales',
suggérant ainsi qu'une multiplication intracellulaire
puisse s'effectuer (Taylor et Brakemore, 1971; Glock
m 1 III .D1SL Il ! ,
.
f f ,.
f
"l
1j
lt
f
" " " ! f ;'<..
t t > ., rt
1(
'ro llJb 15111 2.uma.
tld] . 16 1 •.
.
••
.
"une cong~stion de la muqueuse superfic~él1e,' la n~crose dèS
,
..
cei1ules épithélia1es'et,une forte sédrétion de mucine
prove-, . t I ' 1
nant de l'intérieur des cryptes.
1
, "
Li'étude pathophysiologique
rév~le
.. qUE7' ·la di,arrhéeré-su~te de l'incapacité,des cellules épithéliales du côlon
d'effectuer'l~ transport actif des ions sodium et chlore de
la lumière intestinale vers le sang. On a aussi observé que, . quoique les niveaux d'AMPc et de GMPc soient normaux, leur réponse à un stimulus (théophylline) est fortement atténuée
(Argenzio 'et al., 1980).
Sachant que chez le porc 30 à 50% des sécrétions
endo-"
gènes' sont réab~orbées quotidiennement au niveau du côlon,
il semble très probable qu'une déficience au niveau des mé-canismes d'absorption est suffisante pour expliquer la
déshy-dratatio~ progressive et la mort associées'à la dysenterie
porcine (Argenzi? et a1.( 1980).
Trois mécanismes ont été proposés pour expliquer l'ini-tiaùion de cette colite: l'invasion des cellules épithéliales
par le spirochète, la production 'de ~ytotoxine par le
trépo-nème ou d'autres bactéries anaérobies associées, l'initiation de l'infiltration de la muqueuse par une toxine provenant du
tr~ponème ou d'une autre bactériè (Taylor et Blakemore, 1971;
,
.
/ .:---11 fP.
" ." ..
' j. 1 9 ~'.J.~ ... "u"
(
,
.
, '
" .
Gloék et Harris, 1972; Kent 'et Moon, 1973; Hughes et al.,
1975; Wilcock et 01an~er, 1979). Il n'y'~<cependant aucune
êvidence ferme support~nt l'un ou l'autre de ces mécanismes.
'"
L'hypothèse voulant que l'invasion des cellules
~pi-,
.
tpê~iales par le tréponème soit le mécanisme initi~teur de'
,
la dysenterie porcine est basé~ sur la d~monsttation de la
'.... ,
prêsence de spiroch~tes à l'intérie~ de cellules
épithé-~"'-! •
.
"
'~.lia1es du côlon en dégénérescence (Taylor et Blakemore, 1971; Glock et Harris; 1972). Cependant on n'a pas encore démontré que l'invasion des cel141es:épithé1iales était un prérequis
à l'apparition des lésions (Gloak et al., 1974).
1
, •• • l i ·
L'hypothèse impliquant la production de cypotoxine par le tréponème ou d'autres bactéries anaérobies associées est basée sur l'apparition soudaine de nécrose coagulative de la muqueuse dans les zones où l'on retrouve une grande prolifération de spirochètes et, est aussi basée sur
l'inha-1
bilité de démontrer l'invasion de cellules saines par les
spirocpète~ ou d'au~res bactéries., On a suggéré que les
zones de n~crose sont restreintes à l' é~i théli~.~superficie1
..
parce que des variations du microenvironnement côlonique
pe~ettaient la présence de T. hyodysenteriae et de sa (ses)
toxine (s) à' de~t concentrations adéquates que dans les zones
J d
environnant l'êpithélium superficiel (Kent et Moon, 1973; Hughes et a1., 1975).
~
~~~".M"".~ ••• (._.~".4: ••• ! ••• a ... ,.S .... , ••••• s'.Jia.U.I . . . . I . . . . dt~2.; . . . . b.H1.I.L..4_ •• ta.IU.X~# •• jj~:I.L& . . 4.a.J . . . l.a.a.IINa . . $M[~$ ___ , ~, _______ • _ _ _ " __
•
18
.
.
Enfin, le dernier mécanisme voulant q~e l'initiation
de l'infiltration dans la muqueuse soit du.à une,tqxine
repo-.
se sur l'observation fréquente de caillots dan~ les vaissèaux
sanguins de la muqueuse du
c~lon
depor~.atteints
dè-dysenterie. porcine (Kent et Moon, 1973).
'-:;"
Sachant que T. hyodysenteriae produit un facteur hémolytique et que plusieurs auteurs ont établi une
corré-lation ~irecte eQtre la capacit~ hémolytique de
!.
hyodysenteriaee~ son pouvoir entéropathogène (Taylor et B1akemore, 1971;
Meyer et al., 1974ai Meyer ~t al., 1974b; Meyer ~ al., 1975;
Kinyon et al., 1977), il nous apparai~sait pertinent de
.
procéder â l'étude et à la purification du pouvoir h~molyt'ique
de
!.
hyodysenteriae pour éventuellement en caractériser lemode d'action' et préciser son impl~cation dans la pathogénicité
associée à' .la dysenteri,e porcine.
2.2 Les toxines bactériennes
'
-.
Les toxin~s bactériennes· sont des substances élaborées
,
.
par des bactéries en'ëroissance et sont susceptibles d'altérer ou de détruire les cellules normales humaines ou animales avec lesquelles elles viennent en contact. Cette définition re- . groupe une toule de molécules hétérogènes différant dans leurs
'\
localisâtions dans la ce~lule bactérienne, leur.f structures
chimiques, leurs propriétés physico-ch~miques et biologiques.
- !
1 - i