Restauration des tourbières minérotrophes : études
approfondies des communautés végétales
Thèse
Vicky Bérubé
Doctorat en biologie végétale
Philosophiae doctor (Ph. D.)
Québec, Canada
Restauration des tourbières minérotrophes : études
approfondies des communautés végétales
Thèse
Vicky Bérubé
Sous la direction de :
iii
Résumé
Lors de la mise en place d’un projet de restauration, il est impératif de définir un écosystème de référence qui aidera à orienter les actions à poser. La restauration écologique des tourbières minérotrophes (fens) en est à ses débuts au Canada. À court terme, le but est d’introduire des communautés végétales qui ont le potentiel d’accélérer le retour de l’accumulation de la tourbe. Or, les connaissances actuelles des communautés végétales et des processus écosystémiques propres aux fens sont limitées. Cette recherche doctorale a pour objectif de définir un écosystème de référence pour des fens modérément riches à riches du sud-est du Canada et de cibler des communautés végétales préférentielles à l’atteinte des buts de restauration écologique des tourbières. Afin de consolider l’écosystème de référence, trois sources d’informations (temporelle, spatiale et empirique) ont été utilisées. L’analyse paléoécologique nous informe sur la dynamique de développement hydroséral, somme toute stable, d’un écosystème tourbeux sur une période de 8000 ans. Le comblement d’un plan d’eau est à l’origine de la tourbière où se sont succédées des communautés végétales de grandes cypéracées, puis des communautés dominées par les mousses brunes et les cypéracées basses. Les processus autogènes semblent avoir été prédominants dans l’évolution de la tourbière telle qu’elle est aujourd’hui. La caractérisation de treize fens naturels a résulté à l’identification de deux communautés végétales. La première est une communauté de grandes-cypéracées (magnocariçaie) associée à une production de biomasse et à un niveau d’eau élevés. La seconde est une communauté plus diversifiée de Sphagnum(warnstorfii)-Thuja. À l’exception du S. warnstorfii, cette communauté se compose davantage d’espèces généralistes des tourbières boréales qui croissent dans des habitats plus secs ou ombragés tels que les buttes ou les sous-bois. L’étude met aussi en lumière l’importance du Thuja occidentalis dans les fens régionaux. Sa présence, en densité modérée, devrait être favorisée à long terme. Trois fens naturelles ont été sélectionnées pour une étude approfondie des caractéristiques fonctionnelles de production primaire nette (PPN) et de taux de décomposition. La moyenne de PPN incluant toutes les strates de végétation est de 450 g m-2 an-1. Cette étude confirme l’importance des estimations de PPN du compartiment souterrain (45% de la biomasse totale), des bryophytes (21%) et des arbres (16%). Le S. warsntorfii est la mousse la plus productive (140 g m-2 an-1) et celle qui se décompose le plus lentement (valeur annuelle de décomposition exponentielle k de 0,07). La présence du S. warnstorfii contribue à l’augmentation du potentiel d’accumulation de tourbe. Les communautés du biotope des buttes ont également un meilleur potentiel d’accumulation de tourbe que celles des platières. Finalement, dans la partie expérimentale de la thèse, il a été conclu qu’il n’est pas nécessaire d’augmenter la diversité spécifique et structurelle pour accroître les chances de retour de l’accumulation de tourbe. La performance des variables mesurées est plutôt dépendante de l’identité des espèces composant les assemblages. La présence du Myrica gale, du Carex aquatilis ou du S. warnstorfii dans une communauté augmente significativement la production de biomasse sur le terrain. La présence d’une bryophyte augmente le recrutement de nouvelles espèces tandis que le M. gale ou le C. aquatilis le diminue. La complémentarité entre les espèces a été l’effet dominant sur la production de biomasse dans l’expérience terrain, tandis que l’effet de sélection a été observé en serre. On conclut qu’il existe des relations synergiques dans la décomposition de plusieurs litières mises en association. Une production de biomasse plus élevée devra ainsi être ciblée si le taux global de décomposition est prévu d’augmenter. Suite à la synthèse de ces trois chapitres, il est recommandé d’inclure des bryophytes dans les communautés à restaurer, en particulier le S. warnstorfii afin d’accélérer le retour des fonctions dans les tourbières restaurées et le recrutement de nouvelles espèces. Une communauté de grandes cypéracées est considérée lorsque l’introduction de bryophytes est hasardeuse.
iv
Abstract
When setting up a restoration project, it is imperative to define a reference ecosystem. This will determine the actions to be taken. In Canada, the ecological restoration of the minerotrophics peatlands (fens) is at its infancy stage. In the short term, the restoration goal is to introduce plant communities that have the potential to accelerate the return of peat accumulation. However, current knowledge of plant communities and ecosystem processes specific to fen are limited. The aim of this doctoral research is to define a reference ecosystem for moderately rich to rich fens in southeastern Canada and to target preferential plant communities to achieve the ecological restoration goals of peatlands. In order to consolidate the reference ecosystem, three sources of information were used: temporal, spatial, and empirical. Paleoecological analysis shows us the dynamics of the hydroseral development, notably how stable plant communities has been over a period of 8000 years. Terrestrialization was at the origin of the peatland development. Not long after body of water filled, tall cyperaceae community were followed by communities dominated by the brown mosses and the small cyperaceae. Autogenous processes seem to have been predominant in the evolution of the peatland. The characterization of thirteen natural fens has resulted in the grouping of two plant communities. The first is a tall cyperaceae (magnocaricion) community associated with high biomass production and high water level, the second is a more diverse community of Sphagnum (warnstorfii)–Thuja. With the exception of S. warnstorfii, this community is composed primarily of generalist species of boreal peatlands that grow in drier or shaded habitats, such as underwood or hummocks. The study also highlights the importance of Thuja occidentalis in regional fens. Over time and in moderate density, its presence should be promoted. Three natural fens were selected for an in-depth study of the functional characteristics of net primary production (NPP) and decomposition rates. The average production, including all vegetation layers, is 450 g m-2 yr-1. This study confirms the importance of the NPP estimations with regard to belowground (representing 45% of total biomass), bryophytes (21%) and trees (16%), among others. S. warsntorfii is the most productive bryophyte (140 g m-2 yr-1) and decomposes the slowest (annual exponential decomposition value k of 0.07). The presence of S. warnstorfii contributes to the increased potential for peat accumulation. The communities of the hummock biotope also have a better peat accumulation potential than those of lawns. Finally, in the experimental part of the thesis, it was concluded that there is no need to increase specific or structural diversity to increase the chances of return of peat accumulation. The performance of the measured variables is rather dependent on the identity of the species composing the assemblages. In the field, the presence of Myrica gale, Carex aquatilis or Sphagnum warnstorfii, individually or in combination, increases biomass production. The combination of these three species leads to transgressive overyielding for the production of aerial biomass. The presence of a bryophyte increases the recruitment of new species while M. gale or C. aquatilis will decrease it. The complementarity between species was the dominant effect on biomass production in the field experiment, while the selection effect was observed in greenhouses. Moreover, we concluded that there are synergistic relationships in the decomposition of litters when grouped. Higher biomass production should therefore be targeted if the overall rate of decomposition is predicted to increase. Following the synthesis of the three chapters, it is recommended to include bryophytes in the communities to be restored, in particular S. warnstorfii, in order to accelerate the return of functions in the restored peatlands, and the recruitment and establishment of new species. A community of tall cyperaceae could be considered when the introduction of bryophytes is hazardous and where water levels are forecasted to be high. In short, the preferential plant communities targeted in the reference will accelerate the return of the peat accumulation function and the revival of plant diversity.
v
Table des matières
Résumé ... iii
Abstract ... iv
Table des matières ... v
Liste des tableaux ... viii
Liste des figures ... ix
Remerciements ...xii Avant-propos ...xiv CHAPITRE 1 ... 1 1.1 Problématique... 2 1.2 La restauration écologique ... 4 1.3 L’écosystème de référence ... 5
1.4 Les sources d’informations ... 6
1.5 Identifier le milieu d’études : les tourbières minérotrophes modérément riches ... 8
1.6 État des connaissances sur la restauration des fens ... 12
1.7 Le contexte de cette thèse ... 14
1.8 Questions de recherche ... 16 1.9 Références... 17 CHAPITRE 2 ... 22 Résumé ... 23 Abstract ... 24 2.1 Introduction ... 25
2.2 Materials and methods ... 27
Study sites ... 27
Paleoecological analyses ... 30
Natural site surveys... 30
Data processing and statistical analyses ... 32
2.3 Results ... 34
Paleoecological analyses ... 34
Present-day fen surveys ... 41
2.4 Discussion... 44
Relevance of paleoecological analyses ... 44
Usefulness of modern plant surveys ... 46
Application of the results: BSF case-study ... 47
Complementarity of both methods ... 48
2.5 Acknowledgments ... 51
2.6 References... 52
2.7 Appendix ... 59
Appendix 2.7.1 List of species identified during present-day inventories ... 59
Appendix 2.7.2 Complete macrofossil diagrams ... 64
Appendix 2.7.3 Compilation of the environmental variables ... 67
vi
Appendix 2.7.5 General view of the moderately rich fens inventoried ... 72
CHAPITRE 3 ... 75
Résumé ... 76
Abstract ... 77
3.1 Introduction ... 78
3.2 Materials and methods ... 81
Sites description ... 81
Production measurements ... 82
Decomposition ... 87
Vegetation survey ... 88
Peat accumulation rate ... 88
Statistical analysis ... 88
3.3 Results ... 90
Net primary production... 90
Decomposition ... 90
Biotope comparison ... 91
3.4 Discussion... 97
Overall production in relation to other peatland studies ... 97
Contribution of different vegetation groups to NPP ... 98
Relative above- and belowground production ... 99
Trends for biotopes ... 100
Decomposition: differences among vegetation classes ... 101
Production vs decomposition: potential for peat formation ... 102
Implications for restoration... 102
3.5 Conclusions ... 104
3.6 Acknowledgments ... 105
3.7 References... 106
3.8 Annexes ... 112
Annexe 3.8.1 Données de températures et précipitations ... 112
Annexe 3.8.2 Moyenne (écart-type) de la production primaire nette ... 113
CHAPITRE 4 ... 114
Résumé ... 115
Abstract ... 116
4.1 Introduction ... 117
4.2 Méthodologie ... 121
(1) Expérience combinatoire d’assemblage à dix espèces sur le terrain ... 121
(2) Expérience factorielle d’assemblage à trois espèces en serre ... 129
Calcul des effets de la biodiversité ... 130
Statistiques ... 133
4.3 Résultats ... 135
(1) Les effets de la richesse ... 135
(2) Les effets du groupe structural ... 137
(3.1) L’effet des assemblages sur la production et le potentiel d’accumulation de tourbe ... 141
(3.2) Effet des assemblages sur le recrutement en espèces ... 142
(3.3) Effet de la présence d’une espèce dans un assemblage ... 145
vii
(4.2) Les effets de la biodiversité : complémentarité, sélection et surrendement ... 152
4.4 Discussion... 155
Les effets neutres de la richesse et de la structure ... 155
L’identité de l’espèce prime sur la richesse ou la structure ... 156
Les effets de la biodiversité dans un contexte de restauration de fen ... 157
Approches de restauration pour favoriser le potentiel d’accumulation de tourbe et le retour de la végétation ... 159
Comment augmenter le recrutement de nouvelles espèces... 160
Des pistes de solution pour l’introduction des bryophytes sur tourbe nue ... 161
4.5 Conclusions : recommandations pour une restauration écologique de fens ... 163
4.6 Remerciements... 166
4.7 Références... 167
4.8 Annexes ... 173
Annexe 4.8.1 Dispositif expérimental et photos explicatives des expériences d’assemblages 173 Annexe 4.8.2 Résultats de toutes les analyses de variance en modèle mixte ... 175
Annexe 4.8.3 Liste des espèces inventoriées ... 179
Annexe 4.8.4 (a) Partitionnement de la biomasse aérienne ... 183
Annexe 4.8.5 Effet de biodiversité du surrendement ... 184
Annexe 4.8.6 (a) Effets de biodiversité ... 185
Annexe 4.8.7 Moyennes des principales variables environnementales ... 186
CHAPITRE 5 ... 187 Résumé ... 188 Abstract ... 189 5.1 Introduction ... 190 5.2 Méthodologie ... 192 5.3 Résultats ... 194 5.4 Discussion... 196 5.5 Remerciements... 199 5.6 Références... 200 CHAPITRE 6 ... 203
6.1 Retour sur les résultats, contributions et limites ... 204
Études temporelles ... 204
Études spatiales ... 206
Études empiriques ... 207
6.2 Étude du cas de la tourbière de Bic-Saint-Fabien ... 210
viii
Liste des tableaux
Tableau 1.1 Différentes sources d’informations dans un contexte de restauration des tourbières minérotrophes. ... 6 Table 2.1 Description of 23 environmental variables, code names and units measured in each of the
fens (units) surveyed in the Bas-Saint-Laurent region in 2012. ... 31 Table 2.2 Radiocarbon dates from the four peat cores extracted at the Bic-Saint-Fabien (BSF)
peatland. ... 35 Table 3.1 General description of the three sampled peatlands, all located within 20 km radius. ... 81 Table 3.2 Summary of the methods used for net primary production (NPP) measurements for above
and belowground biomass ... 84 Table 3.3 Mean (±SD) of above- and belowground net primary production (NPP) presented
according to each vegetation class estimated in three moderately rich fens. ... 92 Table 3.4 Summary of vegetation survey done in July 2012 using line-point intercept method in three
moderately rich fens. ... 93 Table 3.5 Summary of net primary production (NPP) for bryophytes and herbs and shrubs groups
(±SD) estimated for lawns and hummocks according to their frequency from vegetation survey observed in each permanent plot. ... 94 Tableau 4.1 Résultats d’analyses chimiques de l’eau et de la tourbe de chaque bloc de l’expérience
combinatoire à Bic-Saint-Fabien. ... 121 Tableau 4.2 Caractéristiques et code de chaque espèce sélectionnée pour l’expérience combinatoire à Bic-Saint-Fabien ... 124 Tableau 4.3 Liste des traitements par assemblage pour l’expérience combinatoire sur le terrain à
Bic-Saint-Fabien ... 126 Tableau 4.4 Liste des traitements par assemblage pour l’expérience factorielle en serre ... 130
ix
Liste des figures
Figure 1.1 Représentation des sources d’information utilisées dans cette thèse pour définir un
écosystème de référence. ... 7 Figure 1.2 Classification des fens selon les gradients de végétation, des fonctions de production et
de décomposition et de chimie de l’eau. Adapté de Hájek et coll. (2006) avec une intégration des résultats de Dierssen et Dierssen (1985 – type de végétation selon le système Braun-Blanquet), Chee et Vitt (1989 – espèces végétales), Zoltaï et Vitt (1995 – gradients et
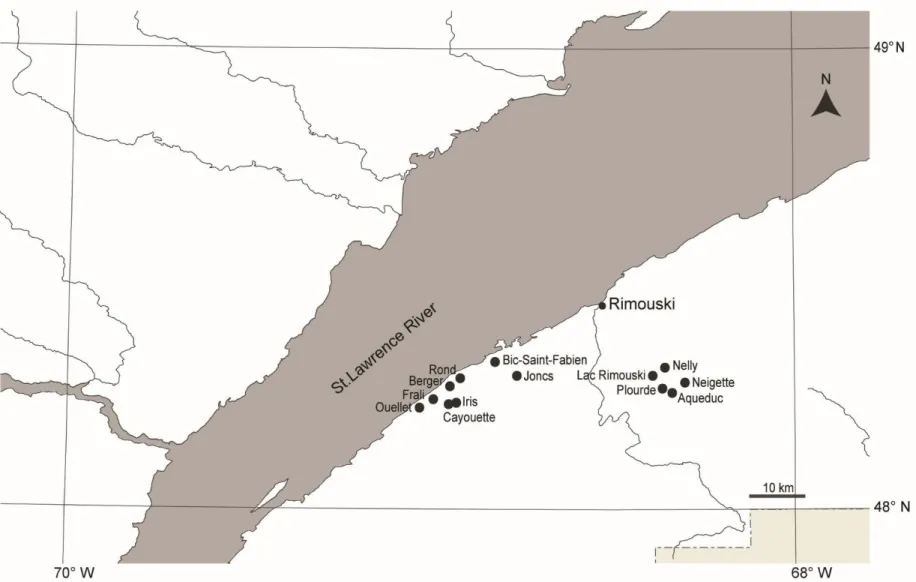
classification), Vitt, Bayley et Jin (1995 – moyenne (écart-type) de la chimie de l’eau de surface, n=18), Vitt (2006 – gradients et classification), Rochefort et coll. (2012 – espèces végétales), Rydin et Jeglum (2013 – espèces végétales, classifications et gradients). ... 11 Figure 2.1 Locations of the 13 sites used for present-day surveys of fen vegetation in the
Bas-Saint-Laurent region, Québec (Canada)... 28 Figure 2.2 Aerial photograph of the Bic-Saint-Fabien (BSF) peatland complex and of the surrounding
landscape. ... 29 Figure 2.3 Macrofossil diagrams (selected taxa) of the Bic-Saint-Fabien BSF peatland. (a) R1, (b) R2, (c) R3, and (d) N. ... 40 Figure 2.4 The 40 most frequent species found in fens of the Bas-Saint-Laurent region, classified by
life form and grouped into two communities obtained by K-means partitioning: (a) tall-sedge and (b) Sphagnum-Thuja communities. ... 42 Figure 2.5 RDA triplot performed with 17 significant environmental variables... 43 Figure 2.6 Schematic representations of the plant assemblages and their associated habitats within a moderately rich fen ecosystem as described by the paleoecological stratigraphies and present-day studies. ... 501 Figure 3.1 (a) Exponential decay coefficient (k yr-1) and (b) percentage of mass loss after 3 years for
bryophytes, Trichophorum cespitosum parts and mix of T. cespitosum and T. nitens. ... 95 Figure 3.2 Average of net primary production (NPP) according to years, months and biotopes (for
each biotope, n=108) for four vegetation classes: (a) graminoids, (b) forbs, (c) ericaceous shrubs and (d) others shrubs species. ... 96 Figure 4.1 Biomasses aériennes, souterraines et totales moyennes estimées sur le (a) terrain et en
(b) serre, ainsi que la décomposition des litières standardisées de (e) racines et de (f) cellulose, du (g) potentiel d’accumulation de la tourbe et du (h) recrutement en nouvelles espèces mis en fonction de la richesse spécifique des assemblages dans l’expérience sur le terrain... 136 Figure 4.2 Résultats de l’expérience sur le terrain à six espèces où la moyenne de la (a) biomasse
x
totale prélevée à la quatrième saison, de la décomposition des litières de (e) racine et de (f) cellulose, du (g) potentiel d’accumulation de la tourbe et du (h) recrutement en nouvelles espèces est mise en fonction des groupes structuraux des assemblages implantés. ... 139 Figure 4.3 Résultats de l’expérience en serre à trois espèces où les moyennes des biomasses (a)
aérienne, (b) souterraine et (c) totale ainsi que (d) le potentiel d’accumulation de tourbe sont mises en fonction des traitements d’assemblages. Chaque assemblage correspond également à un groupe structural défini dans l’expérience terrain. ... 140 Figure 4.4 Moyennes individuelles pour chaque assemblage implanté dans l’expérience de terrain à
six espèces pour les variables de biomasse (a) aérienne prélevées à la deuxième saison, de biomasse (b) aérienne, (c) souterraine et (d) totale prélevées à la quatrième saison, de la décomposition des litières de (e) racine et de (f) cellulose, du (g) potentiel d’accumulation de la tourbe et du (h) recrutement en nouvelles espèces. ... 144 Figure 4.5 Pour l’expérience sur le terrain à six espèces, moyennes de la biomasse aérienne,
souterraine et totale, potentiel d’accumulation de tourbe, recrutement en nouvelles espèces, photo radiation au sol et températures des assemblages avec et sans la présence d’une
espèce dominante établie sur le terrain. ... 147 Figure 4.6 Pour l’expérience sur le terrain à six espèces, valeurs prédites en fonction des valeurs
observées pour le couvert: évolution entre la deuxième, troisième et quatrième saison de croissance. ... 150 Figure 4.7 Valeurs prédites en fonction des valeurs observées pour la biomasse aérienne,
souterraine et totale mesurée dans l’expérience sur le terrain à six espèces (a, b, d, f) et dans l’expérience en serre à trois espèces (c, e, g). ... 151 Figure 4.8 Effets nets de biodiversité (trait noir) décomposée en effet de complémentarité (barres gris
pâles) et effet de sélection (barres gris foncées) estimé à partir (a) de la biomasse aérienne prélevée à la quatrième saison dans l’expérience terrain à six espèces et de la (b) biomasse aérienne et de la (c) biomasse totale prélevées dans l’expérience en serre à trois espèces.. . 154 Figure 5.1 (a) Moyennes du taux linéaire de décomposition k’ mis en fonction de la richesse
spécifique des traitements de litière. (b) Moyenne du taux linéaire de décomposition k’ selon le type de litière estimé en microcosme à toutes les cinq semaines pendant 30 semaines. (c) Les moyennes du taux linéaire de décomposition k’ pour chaque traitement de litière mises en fonction des semaines. Myga = Myrica gale, Caaq = Carex aquatilis et Spwa = Sphagnum warnstorfii. ... 195 Figure 6.1 Résumé des principales conclusions des trois types d’études. ... 209
xi
Pour Danaé, Axel et le petit dernier Et pour moi, parce que j’ai réussi
xii
Remerciements
L’accomplissement de ce projet doctoral a été possible grâce au support et aux encouragements d’une communauté de personne tous plus extraordinaires les unes que les autres. D’abord, un immense merci à Line, ma directrice de thèse, qui m’a accompagnée dans chaque étape de mon travail. Wow que tu as été patiente! J’ai l’impression qu’il y a eu beaucoup d’eau qui a passé sous les ponts depuis mon premier contact avec toi. J’ai eu un parcours atypique, des priorités qui ont changé en cours de route, mais somme toute, je suis parvenue au fil d’arrivée et c’est bien à cause de ta détermination à faire de quoi avec moi! J’ai beaucoup de reconnaissance envers toi : tu es inspirante! Merci!
Jag vill tacka Håkan Rydin (och Uppsala universitet) för ditt varma välkomnade av mig in i er forskningsggrupp. Det var kul att å träffa så munga intressanta människor. Jag uppskattade verkligen alla seminarum, möten och våra diskussioner. Så tack för all din hjälp! Tack så mycket på mina nya kompisar. Lollo och Fia: varje dagar, var solen I Sverige. Kram!
Il y a deux personnes qui ont mis beaucoup d’efforts à m’enseigner à rédiger comme il se doit. Je tiens à remercier Claude Lavoie pour ses (nombreux) commentaires constructifs. Merci également à Chantal Beauchamp qui a lu cette thèse lors du processus de prélecture, qui m’a permis d’améliorer le texte et ma capacité à rédiger de meilleures discussions.
Un énorme merci pour tous les assistants de terrains et de laboratoire qui m’ont aidée sporadiquement ou de façon régulière. Une mention spéciale pour ceux qui m’ont épaulée pendant l’été : Anne-Marie Wagner, Marie-Eve Gauthier, Phan Cat Tuong Le, Catherine Emond, Eugénie Marchand et Kathy Pouliot. Entourée de ces filles sympathiques et motivantes, mes étés ont passé beaucoup trop vite! Je dois un merci spécial à Gilles Berger qui m’a logée pendant mes nombreux déplacements de terrain, qui est devenu un ami et même un assistant de terrain à certaines périodes. Merci à Jean Faubert qui m’a généreusement fait découvrir de beaux coins de sa région qui ont servi à cette thèse. Merci aux propriétaires terriens qui ont accepté que j’inventorie les tourbières qui se trouvaient chez eux. Je dois aussi un remerciement aux autorités du Parc National du Bic (le voisin de mon site d’étude), et ses quelques employés qui m’ont soutenue lors de mes travaux sur le terrain.
xiii
Merci à mes compagnons de bureau; ceux du début (Tommy, Mylène, Josée, Roxane et Myosotis), ceux du milieu (Étienne, Catherine, Virginie et Amélie, Julie), ceux de la fin (Mélina), ceux d’outre-mer (Fia et Charles) et ceux qui ont toujours été là (Sandrine et Rémy). Il y a eu cette belle gang de professionnels de recherche qui m’a écouté, guidé et conseillé: merci Claire, Marie-Claire, Stéphanie et Claudia!
Ma famille et mes amis sont aussi passés au travers de cette thèse en me donnant la tape dans le dos qui me manquait parfois. À mes parents qui m’ont toujours soutenue dans mes projets pas toujours sensés, mais qui ont toujours eu confiance en moi. Ce sont eux qui m’ont trimballée dans la forêt québécoise avec les swamps et les mouches. Veux veux pas, vous êtes à l’origine du désir de faire cette thèse! À ma belle-famille, pour qui cette thèse était un concept bien flou (elle est réelle maintenant!) : vous m’avez bien divertie avec votre bonne humeur. Un immense merci à Marianne qui est devenue une bonne oreille ces dernières années et une amie très précieuse.
À Bruno qui est mon partisan de tous les instants. Celui qui a accepté mon doctorat comme on accepte un ami envahissant (pour toutes les fois que tu ne m’as pas laissée franchir la ligne imaginaire du « je lâche tout! » et que tu m’as laissée travailler là-dessus en essayant de barrer l’accès du bureau/chambre/coin de salon aux enfants, merci!). C’est avec toi que je me suis accomplie et que j’ai produit le meilleur de moi-même, trois fois plutôt qu’une.
À mes trois bébés-thèses, Danaé, Axel et Bébé 3, qui font que je suis meilleure chaque jour. Vous êtes des sources inépuisables de procrastination, mais celles qui valent vraiment la peine d’y mettre du sien. Ma magnifique Danaé, mon vieux génie, maman va pouvoir se trouver un « vrai » travail maintenant! Axel, mon monsieur bonne humeur, maman a déjà creusé avec une pelle mécanique et conduit un tracteur : même moi, ça me surprend tout ce qu’on peut faire pendant un doctorat! Et pour Bébé 3, qui m’a fait la surprise de vouloir m’accompagner pour la dernière étape : j’ai bien hâte de te rencontrer! On fait sa route. Ne l’oubliez jamais.
Finalement, ce doctorat n’aurait pas été possible sans la contribution financière de plusieurs organismes. J’ai eu des bourses nominales de la part du CRSNG et du FQRNT. Le CEN m’a accordé une bourse d’échange pour mon stage à l’étranger. Puis la Chaire de recherche industrielle du CRSNG en aménagement des tourbières a couvert les dépenses logistiques, les frais d’analyses en laboratoire ainsi qu’un financement personnel.
xiv
Avant-propos
L’organisation de cette thèse inclut six chapitres, dont l’introduction générale (chapitre 1) et la conclusion (chapitre 6). Les quatre autres chapitres sont écrits sous forme d’articles scientifiques. Je suis l’auteure principale de ces trois articles. J’ai élaboré les objectifs de recherche, conçu la méthodologie, récolté les données, effectué les analyses statistiques et rédigé les manuscrits. Les trois articles ont été faits en étroite collaboration avec ma directrice de thèse, Line Rochefort. Cette dernière, en tant que coauteure, a pris part à toutes les étapes de production de la thèse et a commenté les manuscrits. Claude Lavoie est aussi coauteur du chapitre 2. Il a collaboré ardemment à la réalisation du volet paléoécologie ainsi qu’à la rédaction de l’article. Le chapitre 2 est accepté officiellement par la revue scientifique Botany et devrait être publié au cours de l’été 2017. Le chapitre 3 a été accepté avec révision à la revue Ecological Indicators. Un manuscrit révisé a été soumis et est en attente.
Chapitre 2 : Bérubé, V., Rochefort, L., et Lavoie, C. Accepté. Fen restoration: defining a reference ecosystem using paleoecological stratigraphy and present-day inventories. Botany. doi: 10.1139/cjb-2016-0281
Chapitre 3: Bérubé, V., et Rochefort, L. Accepté avec révisions. Production and decompositions rates of different fen species as targets for restoration. Ecological Indicators
Chapitre 4 : Bérubé, V., et Rochefort, L. L’impact de la diversité lors de l’établissement d’assemblages végétaux d’une tourbière minérotrophe
Chapitre 5 : Bérubé, V., et Rochefort, L. Décomposition : l’impact de la diversité des litières dans les tourbières minérotrophes
J’ai bénéficié du soutient financier du Conseil de recherches en sciences naturelles et en génie (CRSNG) et du Fonds québécois de la recherche sur la nature et les technologies (FQRNT). Cette thèse a aussi reçu le soutient financier de la Chaire de recherche industrielle du CRSNG en aménagement des tourbières et du ministère des Transports du Québec qui ont couvert les coûts liés aux recherches de terrain et en laboratoire. Le Centre d’études nordiques m’a accordé une bourse d’études pendant mon séjour d’études à l’université d’Uppsala, en Suède. Le département de Växtekologi och evolution du Evolutionsbiologisk centrum de l’Université d’Uppsala et Håkan Rydin
xv
m’ont aussi accordé gracieusement un espace de travail et un soutien pendant la dernière année de cette thèse.
Les résultats de cette thèse ont été présentés lors de diverses rencontres :
- 23e colloque du Groupe de recherche en écologie des tourbières (GRET), Université Laval, Québec, Québec, 21 février 2017
- 81e congrès de l’Acfas – Savoirs sans frontières, Université Laval, Québec, QC, 6-10 mai 2013
- 19e colloque du Groupe de recherche en écologie des tourbières (GRET), Université Laval, Québec, Québec, 21 février 2013 (prix pour meilleure présentation)
- International Symposium on Responsible Peatlands Management and Growing Media Production, Québec, QC, 13-17 juin 2011
- 2e Colloque au Parc national du Bic, Bic, Québec. 7 mai 2011
- 10e Atelier de transfert technologique sur la restauration des tourbières : la remise en eau et la diversité en restauration, Shippagan, Nouveau-Brunswick. 4 et 5 novembre 2010
- Mire Ecosystem Dynamics and Conservation of Biodiversity, Goniadz, Pologne, 16-17 juin 2010
- Reclamation and Restoration of Boreal Peatland and Forest Ecosystems : Toward a Sustainable Future, Edmonton, AB, Canada, 26-27 mars 2010
- 16e colloque du Groupe de recherche en écologie des tourbières (GRET), Université Laval, Québec, Québec. 16 février 2010
- Mini colloque du Centre d’études nordiques, Université Laval, Québec, Québec, 26 février 2009. * prix pour meilleure affiche
1
CHAPITRE 1
2
1.1 Problématique
Une tourbière est un milieu saturé en eau où la production de biomasse est supérieure au taux de décomposition. En conséquence, de la matière organique à forte teneur en carbone, appelée tourbe, s’accumule au fil du temps, d’où l’importance des tourbières dans le cycle mondial du carbone (Gorham 1991; Vasander et Kettunen 2006). Ainsi, les perturbations dans les tourbières sont encore aujourd’hui un sujet préoccupant vu les changements climatiques auxquels la planète fait face. Mondialement, c’est 16 % des tourbières (plus de 487 000 km2) qui ont été dégradées ou détruites par les activités anthropiques (Joosten et Clarke 2002). Au Canada, la situation demeure préoccupante surtout dans la partie méridionale où se concentre la population. Les activités agricoles, forestières, de développement urbain et les activités industrielles d’extraction de tourbe éliminent peu à peu les écosystèmes tourbeux du paysage méridional (Poulin et Pellerin 2001; Pellerin et Poulin 2013). Dans la province du Québec, plus précisément dans la région des basses terres du Saint-Laurent, la superficie des milieux humides, dont 85 % est occupée par des tourbières, a diminué de 19 % sur une période de 22 ans (Pellerin et Poulin 2013). Les mesures de conservation, de compensation, de minimisation ou d’évitement sont, somme toute, peu nombreuses malgré les lois et les politiques mises en place (Joosten et Clarke 2002; Pellerin et Poulin 2013). La restauration est une option de plus qui joue un rôle dans la préservation de l’écosystème des tourbières.
La restauration des tourbières minérotrophes (fens) en est à ses débuts au Canada (Chimner et coll. 2017). Au-delà des techniques de restauration, dans la mise en place d’un projet de restauration, il est impératif de définir une référence (Society for Ecological Restoration International Science and Policy Working Group, SERI 2004; Clewell et Aronson 2013; Shackelford et al. 2013). La référence sert de base pour la définition des objectifs, favorise le choix d’une technique de restauration appropriée et fait office de comparaison lors des suivis post restauration. Dans le cas des tourbières minérotrophes, les connaissances actuelles sur les communautés végétales et les procédés écosystémiques sont surtout régionales. De nombreux inventaires et caractérisations des tourbières dans l’Ouest canadien ont été publiés (ex. Vitt 1990; Gignac 1992; Szumigalski et Bayley 1996; Thormann et coll. 1999; Bayley et Muwort 2004; Locky et coll. 2005) ainsi que la partie arctique et subarctique du Québec (ex. Gauthier et Grandner 1975; Bartsch et Moore 1985). Il y a cependant un manque de connaissances dans la partie méridionale du Canada, soit dans les basses-terres du Saint-Laurent et les Appalaches situées dans le sud-est. C’est pourtant à cet endroit que les besoins de restauration sont en demande.
3
Le but général de restauration des tourbières est axé sur le retour d’un système accumulateur de tourbe (Gorham et Rochefort 2003; Rochefort et coll. 2003; Lamers et coll. 2015; Chimner et coll. 2017). Devant l’étendue de variabilité dans les fens au niveau communautaire et structurel, il serait utile de déterminer la ou les communautés végétales les plus prometteuses à atteindre l’objectif de fonction d’accumulation de carbone. Il est donc essentiel d’obtenir des connaissances telles la composition en espèces et les conditions environnementales associées aux fens modérément riches afin de jeter les bases d’un écosystème de référence en vue d’une restauration écologique. Ce besoin de définir un écosystème de référence avec le choix d'indicateurs devient important vu la diversification des sources d'information et où diverses variables sont proposées. En plus d’une étude historique et de caractérisation de sites naturels, cette thèse propose une approche expérimentale de manipulation de diversité afin de compléter l’information de la référence et ainsi améliorer les résultats d’une restauration écologique de fen.
4
1.2 La restauration écologique
La définition officielle adoptée par le Society for Ecological Restoration International Science and Policy Working Group (SERI 2004) de la restauration écologique est « le processus qui assiste l’autoréparation d’un écosystème qui a été dégradé, endommagé ou détruit ». En d’autres mots, la restauration tente de remettre en place un système autosuffisant et résilient ayant une composition en espèces, une structure et des fonctions similaires à une référence non perturbée. Ce système restauré doit également s’intégrer dans le paysage et prendre en compte des éléments anthropogénies s’il y a lieu, comme l’aspect social et culturel en lien avec les communautés (SERI 2004; Shackelford et coll. 2013).
Un projet de restauration se divise en trois grandes étapes : la planification, la mise en place et le suivi. Après avoir établi la vision que prendra l’écosystème restauré, la planification peut débuter. Elle comprend le choix d’un écosystème de référence, l’identification des besoins de restauration, la définition de buts et la sélection des paramètres pour le suivi. La restauration proprement dite peut ensuite être exécutée en appliquant la ou les techniques appropriées. Les chercheurs se consacrent de plus en plus à vérifier selon la méthode scientifique si certaines techniques se démarquent, en adaptant ou développant des modèles écologiques. La restauration se base ainsi sur la science. Des suivis à court et, si possible, à long terme à l’aide d’indicateurs appropriés sont exécutés afin d’estimer si l’écosystème a pris une trajectoire désirée (Hobbs and Norton 1996). Ce projet de doctorat se concentre essentiellement sur la première étape de la restauration : la définition d’un écosystème de référence.
5
1.3 L’écosystème de référence
Peu importe les objectifs poursuivis, la première étape d’un projet de restauration est de décrire l’écosystème de référence. La référence tient lieu de modèle et servira tout au long du projet. Elle aidera à définir les buts précis de la restauration et ses indicateurs de succès, de même qu’à choisir les techniques de restauration appropriées (White et Walker 1997; Ehrenfeld et Toth 1997; Ehrenfeld 2000; Clewell et Aronson 2013). Elle aide les gestionnaires à prédire comment le système peut devenir autosuffisant (White et Walker 1997). Il est à noter que la référence n’est pas une finalité. Parce qu’il est difficile de prévoir exactement l’évolution de l’écosystème restauré et, qu’à la base, un écosystème est une entité dynamique, il est préférable de voir le projet avec ouverture. Parfois, il est souhaitable de modifier les indicateurs établis au départ et donc la référence.
Plusieurs informations devraient être incluses dans la référence. Principalement, la référence inclut la composition en espèces, la structure et les conditions abiotiques. Les procédés écologiques clés de l’écosystème comme la production primaire, la décomposition ou un cycle biogéochimique particulier comme l’azote doivent aussi être étudiés si l’on désire un retour des interactions dynamiques entre les composantes du système (Martinez 1996; Ruiz-Jaén et coll. 2005). De plus, la restauration vise souvent la continuité historique d’un écosystème sans avoir nécessairement un retour à l’écosystème identique avant les perturbations. La trajectoire historique dicte l’évolution de différents biotopes d’un écosystème à travers le temps. La référence aide à définir les limites de ces écarts et dicte la structure communautaire appropriée (SERI 2004). Des exemples de questions à poser : Est-ce un écosystème avec de longues périodes de stabilité ou en constante évolution? Quelles sont les successions de communautés végétales? Y a-t-il un climax? Le retour de l’écosystème perturbé dans cette trajectoire historique est-il possible? Est-ce que le système restauré peut englober des écarts de variabilité plus ou moins larges?
6
1.4 Les sources d’informations
La référence se base souvent sur une source d’information dite primaire qui provient : (1) soit d’un ou plusieurs des sites naturels et non perturbés situés en périphérie (2) soit sur une partie de l’écosystème qui n’a pas été perturbé (3) ou alors d’une combinaison des deux. Une source d’information secondaire est parfois incluse et consiste à une description de l’écosystème avant la perturbation (Clewell et Aronson 2013).
White et Walker (1997) ont séparé les sources d’informations sur des bases temporelles et spatiales (Tableau 1.1). Or, les données sur les variations temporelles et spatiales sont souvent fragmentaires. Le potentiel de la restauration, l’établissement des buts et l’élaboration des méthodes se basent néanmoins sur ces informations incomplètes. Il peut s’avérer bénéfique d’intégrer plusieurs sources d’informations pour déterminer la référence de façon la plus complète.
Il est possible que des informations concernant les conditions avant les perturbations du site à être restauré soient suffisantes pour bien définir la référence (autoréférence; même place/même temps) ou bien que la perturbation n’ait affecté qu’une partie de l’écosystème (la partie non perturbée est alors considérée comme un refuge; différente place/même temps). Des sites jugés similaires situés à proximité peuvent aussi être utilisés. Afin d’augmenter la validité de la référence, il est possible de se référer à des éléments historiques comme des photographies, des descriptions historiques ou des analyses paléoécologiques du site à être restauré (même place/différent temps) ou, si impossible, à des sites similaires situés à proximité (différente place/différent temps).
Tableau 1.1 Différentes sources d’informations dans un contexte de restauration des tourbières
minérotrophes. Une source d’information primaire inclut les points 1 et 3 tandis qu’une source d’information secondaire inclut les points 2.1, 2.2 et 4. Adapté de White and Walker (1997) et Graf et Rochefort (2008a).
Échelle spatiale
Même lieu Différent lieu
Éc he lle te m pore
lle Même temps 1. Échantillonnage de végétation
avant perturbation fens non perturbés situés à proximité 3. Échantillonnage de végétation de
Différent temps
2.1 Analyse paléoécologique du site perturbé
2.2 Écrits et photographies aériennes historiques
4. Analyse paléoécologique de fens non perturbés situés à proximité
7
Quelques éléments supplémentaires doivent être inclus dans la référence. Dans la mesure du possible, la collecte d’information devrait chercher des études scientifiques de l’écosystème à restaurer, des documents historiques (ex. : photo, cartes, écrits), la mention d’une présence possible d’espèces indésirables (ex. : espèces nuisibles ou envahissantes), les éléments critiques (ex. : espèces clés et cryptiques, moteurs et stress environnementaux) et les éléments manquants (ex. : espèces disparues ou rares). Une attention particulière devrait tenir compte de ces éléments.
Pour cette thèse, nous désirons bâtir concrètement une référence en utilisant des sources d’informations spatiale et temporelle. Cependant, nous ajouterons une troisième source d’information d’origine expérimentale (Figure 1.1). Quoique l’écologie de la restauration se base sur la science, la source d’information expérimentale n’est pas incluse directement dans la définition de la référence. Par le biais de l’approche de la biodiversité fonctionnelle, nous souhaitons déterminer la (ou les) communauté végétale la plus avantageuse pour restaurer un écosystème de tourbières minérotrophes modérément riche à riche.
Figure 1.1 Représentation des sources d’information utilisées dans cette thèse pour définir un
écosystème de référence. L’information temporelle vise à définir une trajectoire historique que le système restauré devrait suivre. L’information spatiale vise à couvrir un certain nombre de sites similaires afin d’estimer une étendue de variabilité de différentes variables (composition en espèce, structures, procédés, variables abiotiques). Pour être considéré sur la bonne trajectoire, le système restauré devrait se situer à l’intérieur de ces limites. Finalement, l’information tirée d’une expérience apporte une réponse à une question spécifique et peut améliorer le recouvrement vers un système résilient et fonctionnel.
8
1.5 Identifier le milieu d’études : les tourbières minérotrophes modérément
riches
Ce projet doctoral se concentre sur un type de tourbière en particulier : les fens modérément riches à riches. Puisqu’il existe de nombreux systèmes de classification des fens, cela laisse croire que les types de fens sont nombreux. Afin éviter la confusion et de mieux situer cette thèse, je présente ici un rapide survol des différents types de fens (Figure 1.2).
La plupart des systèmes de classifications des tourbières se basent sur l’hydrologie, la topographie, la chimie de l’eau ou du sol (pH/l’alcalinité, concentration en nutriment) et l’ontogénie (Figure 1.2). Le système canadien de classification des terres humides (Groupe de travail national sur les terres humides 1997) utilise un système hiérarchique, qui décrit :
(1) la classe (ex. fen, bog, marais) basée sur des critères généraux de végétation, de sol et d’hydrologie;
(2) la forme qui considère le système hydrologique associé à la géomorphologie et la morphologie de surface (terrigène, littogène, soligène ou ombrogène), le régime (riparien, déversement d’eau de surface ou souterraine) et la forme hydrologique (ex. riveraine, lacustrienne, isolée, bassin d’écoulement d’eau douce), et, finalement;
(3) le type basé surtout sur la physionomie des communautés végétales (ex. boisé, arbustif, graminoïde, mousse).
Les fens de bassin arbustif bas, boisé résineux (conifères) et mousses ainsi que les fens ripariens à graminoïdes sont communs dans la région du Bas-St-Laurent et des Appalaches. Cependant, le système canadien ne considère pas la chimie de l’eau, un élément pourtant variable et distinctif.
D’autres classifications des fens se basent selon un gradient de pH et/ou d’alcalinité (disponibilité des cations – surtout Ca2+) et leur végétation associée (Figure 1.2). Au fil du temps, un vocabulaire diversifié et semant parfois la confusion est apparu. Parmi les noms les plus utilisés, en ordre décroissant de pH et d’alcalinité, il y a les fens calcaires (calcareous), les fens très riches (extremely rich), les fens riches (rich), les fens modérément riche (moderately rich), les fens intermédiaires (intermediate), les fens de transition (transitional) et les fens pauvres (poor) (Sjörs 1950; Brighman et
9
coll. 1996; Chee et Vitt 1989; Zolatï et Vitt 1995; Sjörs et Gunnarson 2002; Vitt 2006; Hájek et coll. 2006). Des chevauchements dans la chimie de l’eau et des communautés végétales existent entre les différents types (Sjörs et Gunnarson 2002). Des communautés végétales considérées comme indicatrices d’un type particulier peuvent se trouver dans un autre type de fens dans une autre région (Gore 1983; Glaser 1987), rendant difficile la généralisation ou la rigidité des classifications. Certains auteurs ont simplifié au maximum la classification comme, par exemple, Wheeler et Proctor (2000), qui catégorise tous les fens ayant un pH>5,5 comme fens riches sans considérer les variations des biotopes. Vitt (2006) utilise plutôt trois grandes catégories : pauvre, modérément riche et extrêmement riche. Dans cette classification, les fens modérément riches se confondent aux fens riches décrits par Hájek et coll. (2006) et Sjörs (1950). Cette thèse englobe les catégories de fens modérément riches à extrêmement riches telles que décrites dans Vitt (2006) et révisée dans Rochefort et coll. (2012).
11
Figure 2.1 Classification des fens selon les gradients de végétation, des fonctions de production et de
décomposition et de chimie de l’eau. Adapté de Hájek et coll. (2006) avec une intégration des résultats de Dierssen et Dierssen (1985 – type de végétation selon le système Braun-Blanquet), Chee et Vitt (1989 – espèces végétales), Zoltaï et Vitt (1995 – gradients et classification), Vitt, Bayley et Jin (1995 – moyenne (écart-type) de la chimie de l’eau de surface, n=18), Vitt (2006 – gradients et classification), Rochefort et coll. (2012 – espèces végétales), Rydin et Jeglum (2013 – espèces végétales, classifications et gradients). Les fens de transitions et intermédiaires sont utilisés dans les classifications de fens en Scandinavie, tandis que seulement les fens pauvres, modérément riches et extrêmement riches sont considérés selon Vitt (2006). Plus la teneur en nutriments est élevée, plus la végétation des fens sera dominée par de grandes cypéracées (Kotowski et coll. 2006) jusqu’à être considéré comme un marais (Bayley et Meworth 2004). La structure végétale des fens est régulée par la profondeur de la nappe phréatique : plus celle-ci est basse, comme dans le cas des buttes de mousses, plus les arbres et arbustes seront présents tandis qu’un niveau d’eau près de la surface favorise les cypéracées et mousses brunes (Groupe de travail national sur les terres humides 1997, Vitt 2000).
12
1.6 État des connaissances sur la restauration des fens
L’intérêt grandissant pour la restauration écologique est récent (fin des années 1980). Par exemple, après le constat alarmant que plus de 50 % des milieux humides aux États-Unis ont été drainés ou détruits, le gouvernement américain tente maintenant d’empêcher une plus grande perte des milieux humides et encourage leur restauration (Dahl et Allord 1996). Les programmes gérés par U.S. Army Corps of Engineers Emergency, Natural Resources Conservation Service, U.S. Fish and Wildlife Service et les lois comme le Wetland Resources Act, le Clean Water Act, le Farm Bill, ont propulsé les efforts de restauration des marais, marécages, prairies humides et milieu littoraux (Kentula 1996). Malgré tous ces programmes et lois, la restauration des fens demeure marginale et peu de résultats sont disponibles.
Jusqu’à ce jour en Amérique du Nord (États-Unis et Canada), la restauration des fens se concentre dans les sites avec des perturbations d’origine minières (extraction de tourbe et plateformes de forage). Peu de résultats suite à des projets de restauration de fens ont été publiés. Cobbaert et coll. (2004), Graf et Rochefort (2008b) et Gauthier (2014) ont montré des résultats encourageants pour réintroduire une végétation de fen pauvre et modérément riche à petite échelle en utilisant la technique de transfert muscinale. À grande échelle, les résultats après l’utilisation de la technique du transfert muscinale se sont toutefois avérés décevants en raison de la faible reprise de la végétation (Rochefort et coll. 2016). Des essais de prélèvement mécanique dans l’Ouest canadien pour la restauration de plateformes de forage pétrolier laissent aussi croire que l’approche mécanique n’est pas efficace si les propagules de fens modérément riches sont utilisées (Gauthier, 2014). Au Colorado, les plantations de Carex ont donné des résultats positifs en ce qui concerne le développement du couvert végétal (Cooper et MacDonald 2000) et le remouillage s’avère positif pour retrouver la fonction de puits de carbone (Schimelpfenig et coll. 2014). Diverses techniques ont aussi été explorées dans l’est du Canada, dont les plantations, le remouillage et le remodelage de surface, sans toutefois obtenir des résultats positifs (Rochefort et coll. 2015; Rochefort et coll. 2016).
La situation est différente en Europe. Plusieurs recherches (p. ex., les revues de littératures de Lamers et coll. (2002) et Lamers et coll. (2015)) ont permis de développer des techniques de restauration adaptées aux types de perturbation et à leurs buts. Par exemple, l’utilisation historique des tourbières européennes comme écosystème semi-naturel, c’est-à-dire où l’homme, par son implication, a rendu
13
l’écosystème dépendant de ses intervention les communautés végétales, oriente les interventions vers le retour des pâturages et du fauchage plutôt qu’un retour à un état naturel qui n’existe souvent plus depuis fort longtemps (Klimkowska et coll. 2010; Lamers et coll. 2015). L’eutrophisation des eaux de ruissellement est une autre problématique présente en Europe. Des techniques comme l’enlèvement des couches de surface et le transfert de la paille de fens sont propres aux projets de restauration de fens européens (Klimkowska et coll. 2007). L’expérience européenne s’avère utile pour développer les projets de restauration en développement, mais les différences entre les conditions abiotiques des sites perturbés et les buts de restauration doivent être aussi pris en compte.
14
1.7 Le contexte de cette thèse
Nous croyons que les méthodes et/ou les résultats de cette thèse pourront possiblement s’appliquer lors d’un projet de restauration de fen et ce, peu importe l’étendue de la perturbation. Quoi qu'il en soit, nous croyons qu’il est important de bien situer le projet et le type de perturbation qui est à l’origine de cette thèse.
Cette thèse s’articule autour du projet de restauration de la tourbière de Bic-Saint-Fabien, QC. Cette tourbière, exploitée depuis 1946 par l’industrie de la tourbe, est abandonnée depuis le début des années 2000. La méthode moderne de récolte de la tourbe consiste à retirer la partie vivante de la tourbière, l’acrotelme, à drainer le terrain et aspirer la tourbe par le passage répété de machinerie lourde une mince couche à la fois et ce, aussi longtemps que sa qualité et les conditions de terrain le permettent (Keys 1992). De plus en plus, les tourbières sont utilisées à leur maximum puisque de nouveaux produits incluent de la tourbe de moindre qualité, souvent extraite dans les couches plus profondes. De plus en plus, les compagnies retirent la totalité de la tourbe de sphaigne, exposant ainsi la tourbe de fen sous-jacente. La composition botanique de la tourbe de fen se caractérise par les mousses brunes ou des strates de cypéracées. Les sites exposant la tourbe de fen possèdent des propriétés physico-chimiques propres aux tourbières minérotrophes (Mulder et coll. 1996; Wind-Mulder et Vitt 2000; Graf et coll. 2008) ce qui laisse supposer qu’il serait mieux de restaurer le milieu après l’abandon des activités d’extraction avec une végétation de type minérotrophe, a priori plus apte à coloniser la tourbe résiduelle qui est laissée sur place.
Les conditions hydrologiques et microclimatiques des tourbières abandonnées après l’aspiration de la tourbe empêchent les tapis de mousse de s’établir (Price, 1996). Un réseau de canaux de drainage à chaque 30 mètres abaisse le niveau d’eau (Key 1992), combiné à la compaction et l’oxydation de la tourbe (Schothorst 1977) modifient les propriétés physiques du sol et résultent en une forte variation du niveau d’eau (Schouwenaars 1993; Price 1996). Ces altérations hydrologiques, la formation d’une mince croûte biologique à la surface (encroûtement - Quinty et Rochefort 2003) et le soulèvement gélival (Groeneveld et coll. 2005) font obstacle au rétablissement d’un couvert végétal typique des tourbières. L’intervention active est nécessaire pour rétablir la composition et la structure en espèce à court terme et la fonction d’accumulation de tourbe de l’écosystème à long terme (Gorham et Rochefort 2003; Waddington et coll. 2009). Et avant d’intervenir, il est impératif de bien sélectionner des
15
indicateurs de qualité pour mieux définir la référence, une étape qui n’a pas encore été accomplie pour le projet de restauration de fen de Bic-Saint-Fabien.
16
1.8 Questions de recherche
Cette thèse vise à répondre à cette question : lors d’une restauration écologique de tourbière minérotrophe, quelle(s) communauté(s) végétale(s) doit-on promouvoir afin de ramener l’écosystème vers une trajectoire historique de développement désirée? Cette question en amène une autre : quelle est cette trajectoire historique désirée? Le but général de la thèse est de décrire l’écosystème de référence des tourbières minérotrophes méridionales et émettre des communautés végétales préférentielles. Les attributs de l’écosystème, soit la composition végétale, la structure végétale et les fonctions principales de l’écosystème, seront étudiés en utilisant trois sources d’information, soit en réalisant une étude temporelle, une étude spatiale et étude empirique.
1. Étude temporelle (chapitre 2) : En utilisant la paléoécologie, il sera possible de décrire la trajectoire historique d’une tourbière minérotrophe via ses stades de développement et ses communautés végétales associées. L’étude paléoécologique permettra de déterminer l’importance des bryophytes dans le développement de la tourbière minérotrophe. Le régime naturel des perturbations et le processus de résilience associé aux perturbations seront étudiés. Le stade de régression de la tourbière à restaurer sera aussi évalué.
2. Étude spatiale (chapitres 2 et 3) : En réalisant des inventaires de végétations et une caractérisation des conditions environnementales de sites naturelles, il sera possible de décrire des communautés végétales préférentielles associées à des conditions abiotiques spécifiques. De plus, les deux principales fonctions des tourbières, soit la production de biomasse aérienne et souterraine et le taux de décomposition des principaux groupes de végétation, seront estimées.
3. Étude empirique (chapitres 4 et 5) : La richesse spécifique et structurale sera manipulée dans le but de cibler la communauté végétale ayant le meilleur potentiel d’accumulation de la tourbe. La facilitation entre deux espèces ou plus, et le recrutement de nouvelles espèces sera aussi évaluée. La décomposition de litière mono et plurispécifique sera estimée.
En somme, l’étude temporelle étudie la dynamique de l’écosystème, l’étude spatiale définit concrètement l’étendue de variabilité des composantes de l’écosystème et l’étude empirique cherche à comprendre les effets de la diversité de différentes communautés en établissement afin de maximiser les efforts de restaurations.
17
1.9 Références
Bartsch, L., et Moore, T.R. 1985. A preliminary investigation of primary production and decomposition in four peatlands near Schefferville, Québec. Can. J. Bot. 63(7): 1241-1248. doi:10.1139/b85-171. Bayley, S.E., et Mewhort, R.L. 2004. Plant community structure and functional differences between marshes and fens in the southern boreal region of Alberta, Canada. Wetlands, 24 (2): 277-294. doi:10.1672/0277-5212(2004)024[0277:PCSAFD]2.0.CO;2.
Brighman, S.D., Pastor, J., Janssens, J.A., Chapin, C., et Malterer, T.J. 1996. Multiple limiting gradients in peatlands: a call for a new paradigm. Wetlands, 16: 45-65. doi:10.1007/BF03160645.
Chee, W.-L., et Vitt, D.H. 1989. The vegetation, surface water chemistry and peat chemistry of moderate-rich fens in central Alberta, Canada. Wetlands, 9(2): 227-261. doi:10.1007/BF03160747. Chimner, R.A., Cooper, D.J., Wurster, F.C., et Rochefort, L. 2017. An overview of peatland restoration in North America: where are we after 25 years? Restor. Ecol. 25(2): 283-292. doi: 10.1111/rec.12434 Clewell, A.F., et Aronson, J. 2013. Ecological restoration principles, values, and structures of an emerging profession. Second edition. Island Press, Washington.
Cobbaert, D., Rochefort, L., et Price, J.S. 2004. Experimental restoration of a fen plant community after peat mining. Appl. Veg. Sci. 7: 209-220.
Cooper, D.J., et MacDonald, L. 2000. Restoring the vegetation of mined peatlands in the southern Rocky Mountains of Colorado, USA. Restor. Ecol. 8:103-111. doi:10.1046/j.1526-100x.2000.80016.x. Dahl, T.E., et Allord, G.J. 1996. History of wetlands in the conterminous United States. In National water summary on wetland ressources: Technical aspects. United States geological Survey Water-Supply Paper 2425. Pp. 19-26.
Dierssen, K., et Dierssen, B. 1985. Suggestions for a common approach in phytosociology for Scandinavian and Central European mire ecologists. Aquilo Ser Botanica 32 : 33-44.
Ehrenfeld, J.G. 2000. Defining the limits of restoration: the need for realistic goals. Restor. Ecol. 8: 2-9. doi:10.1046/j.1526-100x.2000.80002.x.
Ehrenfeld, J.G., et Toth, L.A. 1997. Restoration ecology and the ecosystem perspective. Restor. Ecol. 5: 307-317. doi:10.1046/j.1526-100X.1997.00544.x.
Gauthier, M.-E. 2014. Restoring peatland plant communities on mineral well pads. Mémoire de M.Sc., Université Laval, Québec, Québec. 63 p.
Gauthier, R., et Grandtner, M.M. 1975. Études phytosociologique des tourbières du Bas-St-Laurent, Qc. Nat. Can. 102(1): 109-153.
18 Gignac, L.D. 1992. Niche structure, resource partitioning, and species interactions of mire bryophytes relative to climatic and ecological gradients in Western Canada. Bryologist, 95(4): 406-418. doi:10.2307/3243564.
Glaser, P.H. 1987. The development of streamlined bog islands in the continental interior of North America. Arctic Alpine Res. 19(4): 402-413. doi:10.2307/1551405.
Gore, A.J.P. (éd.). 1983. Ecosystems of the world. 4A. Mires : swamps, bog, fen and moor. Regional studies. Elsevier, Amsterdam.
Gorham, E. 1991. Northern peatlands: role in the carbon cycle and probable response to climatic warming. Ecol. Appl. 1:182–195. doi:10.2307/1941811.
Gorham, E., et Rochefort, L. 2003. Peatland restoration: a brief assessment with special reference to Sphagnum bogs. Wetl. Ecol. Manag. 11: 109-119. doi:10.1023/A:1022065723511.
Graf, M.D., Rochefort, L., et Poulin, M. 2008. Spontaneous revegetation of harvested peatlands of Canada and Minnesota, USA. Wetlands, 28(1): 28-39. doi:10.1672/06-136.1.
Graf, M.D., et Rochefort, L. 2008a. Restoring peat-accumulating function on cutaway peatlands. Dans Proceedings of the 13th International Peat Congress: After Wise Use – The Future of Peatlands, Volume 1: Oral Presentations, Tullamore, Ireland, 8 - 13 June 2008. Édité par C. Farrell et J. Feehan. International Peat Society, Jyväskylä, Finlande. Pp. 398- 400.
Graf, M.D., et Rochefort, L. 2008b. Techniques for restoring fen vegetation on cut-away peatlands in North America. Appl. Veg. Sci. 11: 521-528. doi:10.3170/2008-7-18565.
Groeneveld, E.V.G., et Rochefort, L. 2005. Polytrichum strictum as a solution to frost heaving in disturbed ecosystems: a case study with milled peatlands. Restor. Ecol. 13(1): 74-82. doi:10.1111/j.1526-100X.2005.00009.x.
Groupe de travail national sur les terres humides. 1997. Système de classification des terres humides du Canada. 2e édition. Édité par B.G. Warner et C.D.A. Rubec. University of Waterloo, Ontario. Hájek, M., Horsák, M., Hájkova, P., et Dítě, D. 2006. Habitat diversity of central European fens in relation to environmental gradients and an effort to standardize fen terminology in ecological studies. Perspect. Plant Ecol. 8:97-114. http://dx.doi.org/10.1016/j.ppees.2006.08.002.
Hobbs, R.J., et Norton, D.A. 1996. Toward a conceptual framework for restoration ecology. Restor. Ecol. 4: 93-110. doi:10.1111/j.1526-100X.1996.tb00112.x.
Joosten, H., et Clarke, D. 2002. Wise use of mires and peatlands - Background and principles including a framework for decision-making. International Peat Society et International Mire Conservation Group, Finland.
Kentula, M.E. 1996. Wetland restoration and creation. National water summary on Wetland resources: Technical aspects. United States geological Survey Water-Supply Paper 2425. Pp. 87-92.
19 Keys, D. 1992. Canadian peat moss and the environment. Issues Paper, No. 1992–3. North American Wetlands Conservation Council, Canada.
Klimkowska, A., van Diggelen, R., Bakker, J.P., et Grootjans, A.P. 2007. Wet meadow restoration in Western Europe: a quantitative assessment of the effectiveness of several techniques. Biol. Conserv. 140: 318-328. https://doi.org/10.1016/j.biocon.2007.08.024
Klimkowska, A., Grootjans, A.P., Kotowski, W., et van Diggelen, R. 2010. Prospects for fen meadow restoration on severely degraded fens. Perspect. Plant Ecol. 12: 245-255.
https://doi.org/10.1016/j.ppees.2010.02.004
Kotowski, W., Thörig, W., van Diggelen, R., et Wassen, M.J. 2006. Competition as a factor structuring species zonation in riparian fens – a transplantation experiment. J. Appl. Veg. Sci. 9(2): 231-240. http://www.jstor.org/stable/4620482.
Lamers, L.P.M., Smolders, A.J.P., et Roelofs, J. 2002. Restoration of fens in the Netherlands. Hydrobiologia 478: 107-130. doi:10.1023/A:1021022529475.
Lamers, L.P.M., Vile, M.A., Grootjans, A.P., Acreman, M.C., van Diggelen, R., Evans, M.G., Richardson, C.J., Rochefort, L., Kooijman, A.M., Roelofs, J.G.M., et Smolders, A.J.P. 2015. Ecological restoration of rich fens in Europe and North America: from trial and error to an evidence-based approach. Biol. Rev. 90: 182-203. doi:10.1111/brv.12102.
Locky, D.A., et Bayley, S.E. 2005. Plant diversity, composition, and rarity in the southern boreal peatlands of Manitoba, Canada. Can. J. Bot. 84: 940-955. doi:10.1139/b06-049.
Martinez, N.D. 1996. Defining and measuring functional aspects of biodiversity. Dans Biodiversity: A Biology of Numbers and Difference. Édité par K.J. Gaston. Blackwell, Oxford, UK.
Pellerin, S., et Poulin, M. 2013. Analyse de la situation des milieux humides au Québec et recommandations à des fins de conservation et de gestion durable. Centre de la Science de la Biodiversité du Québec. 104 p. + annexes.
Poulin, M., et Pellerin, S. 2001. La conservation. Dans Écologie des tourbières du Québec-Labrador. Édité par S. Payette et L. Rochefort. Presses de l'Université Laval, Québec, Québec. Pp. 505-518. Price, J.S. 1996. Hydrology and microclimate of partly restored cutover bog, Québec. Hydrol. Process. 10 : 1263-1272. doi:10.1002/(SICI)1099-1085(199610)10:10<1263::AID-HYP458>3.0.CO;2-1. Quinty, F. et Rochefort, L. 2003. Guide de restauration des tourbières, 2e éd. Association canadienne de mousse de sphaigne et Ministère des Ressources naturelles du Nouveau-Brunswick. Québec, Québec.
Rochefort, L., Quinty, F., Campeau, S., Jonhson, K., et Malterer, T. 2003, North American approach to the restoration of Sphagnum dominated peatlands, Wetl. Ecol. Manage. 11(1-2): 3-20. doi:10.1023/A:1022011027946.
20 Rochefort, L., Strack, M., Poulin, M., Price, J.S., Graf, M.D., Desrochers, A., et Lavoie, C. 2012. Northern peatlands. Dans Wetland Habitats of North America: Ecology and Conservation Concerns. Édité par D.P. Batzer et A.H. Baldwin, University of California Press, Berkeley, Californie. Pp. 119-134. Rochefort, L., LeBlanc, M.-C., Pouliot, R., Hogue-Hugron, S., D’Amour, N., et Boismenu, C. 2015. Restauration écologique des tourbières de Bic–Saint-Fabien et de Saint-Fabien-sur-Mer dans le Bas-Saint-Laurent. Projet R719.1, Rapport final présenté au Ministère des Transports du Québec. Groupe de recherche en écologie des tourbières, Université Laval, Québec. 90 p. et 3 annexes.
Rochefort, L., LeBlanc, M.-C., Bérubé, V., Hogue, S., Boudreau, S., et Pouliot, R. 2016. Reintroduction of fen plant communities on a degraded minerotrophic peatland. Botany, 94(11): 1041-1051. doi :10.1139/cjb-2016-0023
Ruiz-Jaén, M.C., et Aide, T.M. 2005. Restoration success: how is it being measured? Restor. Ecol. 13(3): 569-577. doi:10.1111/j.1526-100X.2005.00072.x.
Rydin, H., et Jeglum, J.K. 2013. The Biology of Peatlands. Second edition.: Oxford University Press, New York.
Schimelpfenig, D.W., Cooper, D.J., et Chimner, R.A. 2014. Effectiveness of ditch blockage for restoring hydrology and soil processes in mountain peatlands. Restor. Ecol. 22(2): 257-265. doi:10.1111/rec.12053.
Schothorst, C.J. 1977. Subsidence of low moor peat soil in the western Netherlands. Geoderma, 17: 265-291. doi:10.1016/0016-7061(77)90089-1.
Schouwenaars, J.M. 1993. Hydrological differences between bogs and bog-relicts and consequences for bog restoration. Dans Netherlands-Wetlands. Proceedings of a Symposium held in Arnhem, The Netherlands, December 1989. Édité par E.P.H. Best et J.P, Bakker. Developments in Hydrobiology series vol. 88. Springer Netherlands, Pays-Bas. Pp 217-244. doi:10.1007/978-94-011-2042-5_11. SERI (Society for Ecological Restoration International Science and Policy Working Group). 2004. The SER International Primer on Ecological Restoration. Society for Ecological Restoration International, Tuscon, AZ. [online]. Disponible au www.ser.org.
Shackelford, N., Hobbs, R.J., Burgar, J.M., Erickson, T.E., Fontaine, J.B., Laliberté, E., Ramalho, C.E., Perring, M.P., et Standish, R.J. 2013. Primed for Change: Developing Ecological Restoration for the 21st Century. Restor. Ecol. 21: 297–304. doi:10.1111/rec.12012.
Sjörs, H. 1950. On the relation between vegetation and electrolytes in North Swedish mire waters. Oikos, 2: 241–259. doi:10.2307/3564795.
Sjörs, H., et Gunnarsson, U. 2002. Calcium and pH in north and central Swedish mire waters. J. Ecol. 90(4): 650-657. doi:10.1046/j.1365-2745.2002.00701.x.