Contribution des réserves calciques présynaptiques et gliales dans la modulation de la transmission synaptique à la jonction neuromusculaire de la grenouille Rana pipiens
269
0
0
Texte intégral
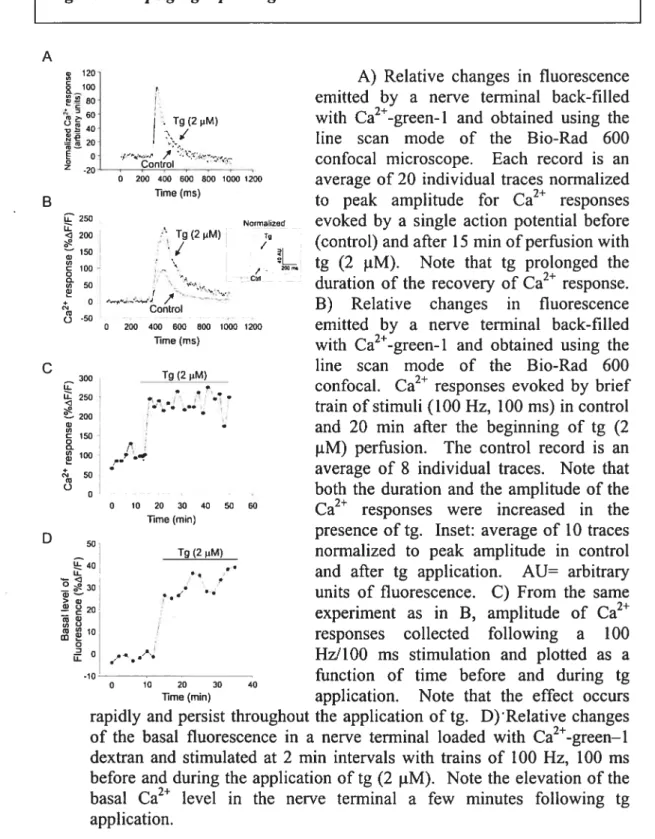
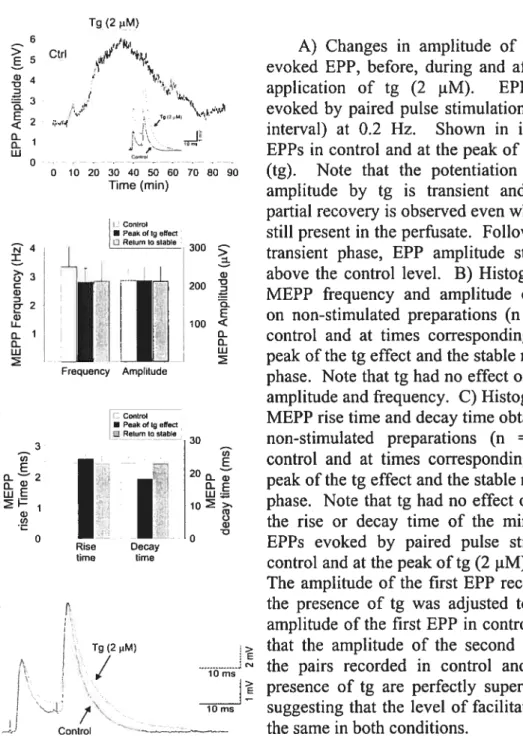
Figure

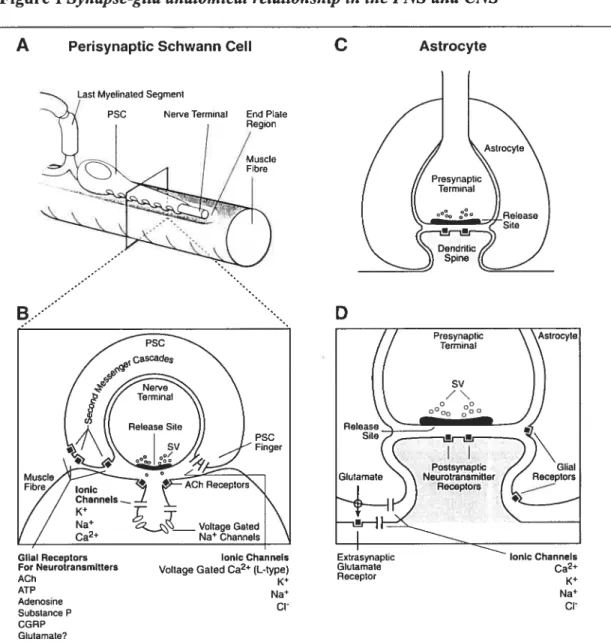
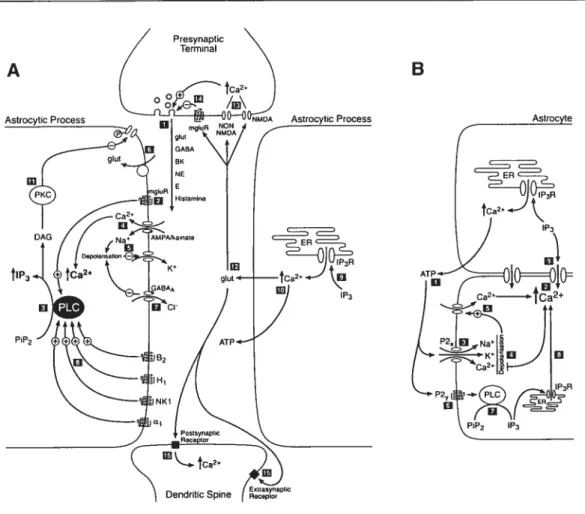

+7
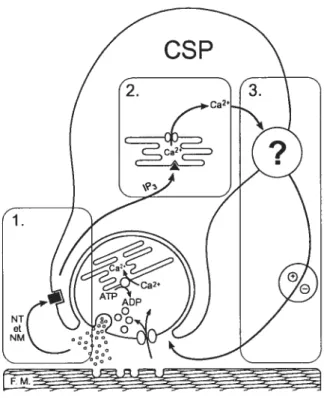

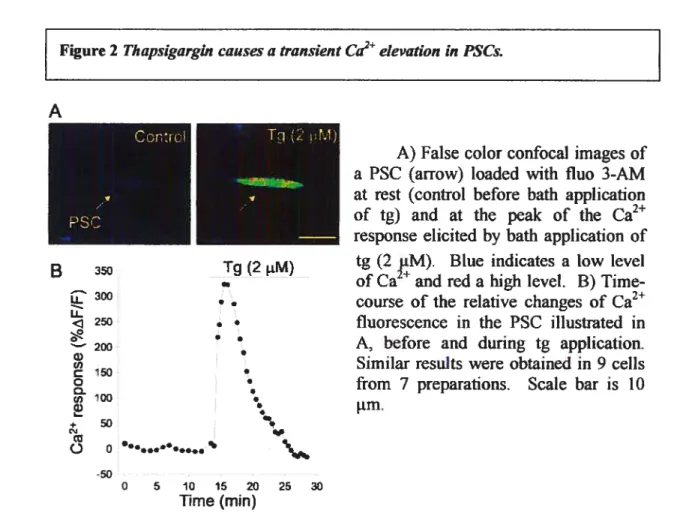

Documents relatifs
