HAL Id: hal-02821616
https://hal.inrae.fr/hal-02821616
Submitted on 6 Jun 2020HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Mesures de l’interception lumineuse et de la
photosynthèse de jeunes hêtres sous couvert de pin dans
la chaîne des Puys
Caroline Allen
To cite this version:
Caroline Allen. Mesures de l’interception lumineuse et de la photosynthèse de jeunes hêtres sous couvert de pin dans la chaîne des Puys. [Stage] Université Blaise Pascal (Clermont Ferrand 2) (UBP), FRA. 2008, 26 p. �hal-02821616�
Caractérisation de l'interception
de la lumière et du
fonctionnement photosynthétique
de jeunes hêtres
Master 1 Génétique et Physiologie Végétale
Membres du Jury: Aurélie Gousset Gérard Ledoigt
Saïd Mouzeyar 03 Septembre 2008
UNIVERSITE BLAISE PASCAL - CLERMONT II -
UFR Sciences Exactes et Naturelles
Centre INRA de Clermont-Ferrand
Site de Crouelle
234 avenue du Brézet
63100 Clermont-Ferrand cedex 02
Maître de stage: Philippe Balandier
Tutrice de stage: Aurélie Gousset
Responsable de stage: Nathalie Fournier
2
Sommaire
Résumé... 3 Abréviations... 4 Introduction... 5 Bibliographie... 4 Matériel et méthodes ... 8 1. Site et matériel... 8 2. Méthodes ... 8 a. Mesures de photosynthèse... 8b. Mesures de l’interception de la lumière ... 9
c. Mesures dendrométriques ...10 Résultats ... 11 Discussion ...12 Conclusion...14 Références bibliographiques...15 Annexes...17
3
Résumé
Le hêtre (Fagus sylvatica) est une espèce peu présente dans les forêts de la Chaîne des Puys. Sa tolérance à l’ombrage est étudiée par l’équipe AMi de l’UMR PIAF ainsi que le CEMAGREF dans le but de le réintroduire sous couvert de pins. Pour cela, l’interception de la lumière et le fonctionnement photosynthétique de jeunes hêtres sont analysés.
Les changements physiologiques liés à la lumière sont mesurés à travers 3 grands paramètres: la photosynthèse, la respiration et la lumière interceptée. Les changements morphologiques sont étudiés par mesures des paramètres dendrométriques des jeunes arbres.
Leur croissance à différents degrés d’éclairement montre une plasticité des traits physiologiques et morphologiques tels que le contenu en azote foliaire ou l’arrangement spatial des feuilles.
Summary
The beech (Fagus sylvatica) is a species not very present in the forests of the Chaîne des Puys. Its tolerance with the shade is studied by the team AMi of UMR-PIAF in cooperation with the CEMAGREF in order to reintroduce it under the covers of Pines. For that, the light interception and the photosynthetic functioning of young beeches are analyzed.
The physiological changes are measured through 3 great parameters: photosynthesis, respiration and intercepted light. The morphological changes are studied by measurements of the dendrometric parameters of the young trees.
Their growth with various degrees of light shows a plasticity of the physiological and morphological treats such as the leaf content nitrogen or leaf spatial arrangement.
4
Abréviations
PARt: Photosynthetic Active Radiation Transmitted LA: Leaf Area
STAR: Silhouette to Total LA Ratio
Rubisco: Ribulose 1, 5 bisphosphate carboxylase/oxygénase Vcmax: Vitesse de carboxylation du RuBP par la Rubisco Jmax : vitesse de transport des électrons
Rd : dark Respiration 3D : tridimensionnel
Na : concentration d’azote par unité de surface de feuille LMA : masse de matière sèche par unité de surface foliaire
5
Introduction
Une grande partie de la Chaîne des Puys était autrefois utilisée à des fins agricoles ou d’élevage (pâturages communaux et zones de cultures). Les deux guerres mondiales de 1914-1918 et 1939-1945 ont provoqué un exode rural vers Clermont-Ferrand entraînant l’abandon de ces terres et, par conséquent, leur enfrichement ou boisement spontané par colonisation d’espèces ligneuses telles que le noisetier (Corylus avellana), le pin sylvestre (Pinus sylvestris) ou le bouleau (Betula pendula).
Le développement de ces nouveaux boisements peu diversifiés et à faible valeur économique ont entraîné la fermeture des paysages et augmenté les risques d’incendie. La hêtraie (Fagus sylvatica) est l’aboutissement de l’évolution de ces boisements; c’est, sous le climat considéré, le stade terminal de la succession écologique des prairies aux forêts (Prévosto et Balandier 2007). C’est actuellement la principale végétation climacique de la Chaîne des Puys. Le hêtre y est donc une espèce recherchée qui apporte une valeur écologique, économique et patrimoniale importante. On cherche donc à favoriser son installation.
Les recherches entreprises par l’équipe AMi de l’UMR-PIAF et le CEMAGREF ont pour objectif d’analyser la croissance de jeunes hêtres en relation avec le PAR disponible sous le couvert de boisements de pin sylvestre et de tester l’hypothèse d’une modification de leurs traits physiologiques et morphologiques associés à la tolérance à l’ombre. Distinguer les effets directement liés à la lumière des effets liés à la dimension des plants est une étape importante de cette étude, qui a fait l’objet de mon stage.
Schéma 1: Influence du temps sur l’évolution de la compétitivité des différents groupes de végétation en milieu forestier.
La compétitivité évolue d’un type de végétation à un autre avec le temps.
4
Bibliographie
La biodiversité des milieux forestiers est la source d’un dynamisme de la végétation et n’est envisageable que par cohabitation de différentes espèces. En forêt, plusieurs étages peuvent se distinguer: l’étage principal ou étage supérieur, le mi-étage et le sous-bois. Les grands groupes d’espèces sont : les arbres d’étage principal, les grands arbustes et les arbres de mi-étage, les fougères et graminées du sous-bois (Balandier, Collet et al. 2006).
Chacun de ces groupes a un effet compétitif sur la végétation qui l’environne pour les ressources naturelles telles que l’eau, la lumière, les éléments minéraux du sol… Ainsi, les graminées forment une barrière à l’eau sur une profondeur d’un mètre. Les milieux clairsemés sont rapidement colonisés par les fougères ce qui réduit fortement la lumière au niveau du sol. Les petits arbustes ont un système racinaire dense et profond ce qui réduit le taux d’éléments nutritifs dans le sol. Les grands arbustes, les arbres de mi-hauteur et les arbres de l’étage supérieur sont compétitifs pour l’interception de la lumière. Les compétiteurs de sous-bois (petits arbustes et fougères principalement) sont préjudiciables la première année après établissement des jeunes plants d’arbre tandis que ceux de mi-étage et étage supérieur sont des compétiteurs principalement pour la lumière (mais restent très variés) et ont des effets à plus long terme. La compétitivité évolue d’un type de végétation à un autre avec le temps (cf. schéma 1) (Balandier, Collet et al. 2006).
Sous le pédoclimat de la Chaîne des Puys, la succession des prairies aux forêts sur des territoires agricoles laissés à l’abandon aboutit théoriquement à la hêtraie. C’est le stade terminal de l’évolution des boisements spontanés (Prévosto et Balandier 2007).
Le hêtre est un arbre d’étage supérieur qui se régénère en sous-bois, donc à faible éclairement. C’est une espèce tolérante à l’ombrage très compétitive pour la lumière. Il investit préférentiellement sa biomasse dans sa surface foliaire plutôt que dans les parties ligneuses ou
5 racinaires contrairement aux essences de lumière. Cet investissement est représenté par le LAR ou Leaf Area Ratio, soit le rapport entre tissus photosynthétiques et tissus non-photosynthétiques. Une étude a montré qu’il est de 50% plus faible pour les grands arbres que pour les petits de façon générale (Balandier, Sinoquet et al. 2007). Le LAR étant un paramètre variable au cours du développement des jeunes hêtres, il est primordial de distinguer les effets de dimensions et les effets de lumière sur leur croissance.
Il a été montré que les traits morphologiques des jeunes arbres sont adaptés aux conditions de lumière. Les angles d’inclinaison des feuilles tendent vers la verticale afin de réduire la photoinhibition en pleine lumière ou vers l’horizontale afin de diminuer l’auto-ombrage en sous-bois (Fleck, Niinemets et al. 2003).
En cas de forte lumière, les bords du limbe des feuilles sont recourbés pour éviter la photoinhibition. Le parenchyme palissadique contenant les enzymes de photosynthèse (limitantes dans ces conditions) est donc très développé ainsi que la masse de matière sèche par unité de surface foliaire. Au contraire, à l’ombre, c’est l’interception de la lumière qui est limitante d’où des feuilles moins épaisses en sous-bois qu’en pleine lumière. De plus, en faible lumière, les feuilles sont planes et leur arrangement spatial limite l’auto-ombrage.
Les adaptations morphologiques tendent à capter le maximum de lumière à l’ombre et les adaptations physiologiques à conserver un gain de carbone net positif en en limitant les pertes. Ainsi le contenu en azote foliaire, les taux de photosynthèse et de respiration et le taux de croissance sont plus élevés à la lumière (Balandier, Collet et al. 2006).
Pour étudier ces paramètres, plusieurs techniques on été mises au point.
La digitalisation 3D permet d’évaluer l’arrangement spatial du feuillage (inclinaison et position des feuilles) et donc l’interception de la lumière par la surface foliaire de l’arbre tandis que des mesures de photosynthèse permettent d’évaluer l’efficacité de fixation du dioxyde de carbone par
6 la Rubisco dans les carbohydrates. La conversion de l’énergie lumineuse en énergie chimique est un moyen d’estimer la croissance du jeune arbre quelle que soit sa taille (Balandier, Sinoquet et al. 2007) et en fonction des conditions d’éclairement. En effet, la croissance demande un apport important de carbone.
Le modèle de Farquhar (Farquhar, Von Caemmerer et al. 1980) est appliqué pour traiter les mesures de photosynthèse. Il s’agit du modèle le plus fréquemment utilisé pour décrire les variations d’assimilation nette en CO2 d’une feuille en fonction des paramètres de l’environnement des plantes en C3 telles que Fagus sylvatica. Il résulte de l’intégration à l’échelle foliaire de caractéristiques mesurables in vitro à l’échelle du chloroplaste (Farquhar, Von Caemmerer et al. 1980). Ce modèle est basé sur le processus de la photosynthèse dans un environnement stable et est centré sur les propriétés cinétiques de la Rubisco, enzyme clé de la photosynthèse. L’assimilation nette de CO2 est considérée comme étant le résultat de trois processus : la carboxylation du Ribulose-1,5-biphosphate(ou RuBP), ce qui consomme du CO2, l’oxygénation du RuBP qui est une réaction compétitive pour le même site actif et qui conduit à la photorespiration (1 mole de CO2 produite pour 2 moles d’O fixées) et enfin, la respiration liée aux phosphorylations oxydatives dans les mitochondries(Rd) produisant du CO2 (Farquhar, Von Caemmerer et al. 1980). La vitesse de carboxylation est conditionnée par la quantité de Rubisco dans la feuille et par ses propriétés cinétiques qui dépendent des concentrations relatives des deux substrats (CO2 et O2), de la concentration de RuBP par rapport à celle du site actif de la Rubisco pour le RuBP, de la concentration en inhibiteur (carboxyarabinitol-1-p) et de l’état d’activation de la Rubisco. Ces deux derniers paramètres ne sont pas pris en compte dans le modèle de Farquhar. Ce modèle nécessite de réaliser les mesures en conditions optimales pour l’activation de la Rubisco. La spécifité de celle-ci pour le CO2 plutôt que pour l’O2 est thermosensible. La température optimale est 25°C. Mais l’activité de la Rubisco peut être saturée en RuBP (son activité est alors contrôlée par ses propriétés cinétiques et les concentrations en CO2 et O2) ou
7 limitée en RuBP (par la régénération de la RuBP). En conditions in vivo, le second cas est très peu fréquemment rencontré, il n’est observé que lorsque le CO2 est en très forte quantité ou en cas de carence de phosphate. La respiration à l’obscurité(Rd) est soit estimée par ajustement sur les courbes de réponse au CO2 sous une lumière forte soit mesurée directement à l’obscurité. Son estimation reste un problème encore non résolu.
Dans la pratique, la capacité de photosynthèse des feuilles est représentée par deux paramètres : Vcmax et Jmax. Le Vcmax est la vitesse de carboxylation par la Rubisco et le Jmax est la vitesse de transport des électrons. Le protocole consiste à établir une courbe de réponse au CO2 sous une lumière constante. Les paramètres cinétiques de la Rubisco sont considérés comme invariables d’une espèce à l’autre et d’un génotype à l’autre (pour les plantes C3). Le modèle de Farquhar a été amélioré par Harley et Tenhunen(1991).
La tolérance à l'ombrage est actuellement définie comme étant la capacité de survie à faible niveau d'éclairement pendant de longues durées et à reprendre rapidement une croissance active après une remise en lumière. La croissance augmente avec la disponibilité en lumière jusqu’à un niveau optimal et diminue en plein découvert. Les tissus non-photosynthétiques se développant plus rapidement que les tissus photosynthétiques, LAR diminue avec la croissance du plant ainsi que l’efficacité d’interception de la lumière. Il faut donc de plus en plus de lumière au fur et mesure qu’il grandit pour conserver un bilan de carbone positif.
Distinguer les effets dus à la lumière des effets dus à la taille des jeunes plants sur leur croissance est une étape essentielle de cette étude. L'hypothèse d'une dimension critique à partir de laquelle le bilan de carbone ne serait plus positif (appareil foliaire insuffisant pour maintenir les tissus non-photosynthétiques) a été proposée. La dimension critique, dans le cas du hêtre serait plus élevée que celle des essences de lumière.
Le suivi de croissance sur plusieurs années va permettre d'étudier ces hypothèses et les confirmer ou non.
Situation géographique des placettes d’expérimentation et photographies des placettes 1, 2, 4 et 7.
1
2
4
8
Matériel et méthodes
1. Site et matériel
Plusieurs placettes situées au sud de la Chaîne des Puys dans la section de Fontfreyde de la commune de St-Genès-Champanelle sur la cheire volcanique de Mercœur ont été aménagées en 2001. Ce dispositif a été élaboré dans le but de créer un gradient d’éclairement en sous-bois par des placettes ayant des couverts de pins plus ou moins denses et âgés.
Sur chacune d’elles ont donc été installés en 2001 de jeunes hêtres caractéristiques des boisements naturels de la zone. La placette témoin(7) est placée en pleine lumière sur une prairie. Les hêtres qui y sont plantés sont désherbés pour éviter la compétition des herbacées pour l’eau et les nutriments du sol. Un suivi in situ de croissance de hêtres est effectué suivant l’éclairement depuis 2001. Pour cette étude, douze hêtres ont été sélectionnés afin d’obtenir un gradient d’éclairement entre les différents arbres et indépendamment de la placette. La digitalisation est réalisée sur une partie de la sélection et des mesures de photosynthèse sont réalisées sur deux feuilles au moins de chacun des hêtres qui sont ensuite collectées afin de réaliser des dosages d’azote et mesures de surface foliaire.
2. Méthodes
a. Mesures de photosynthèse
L’appareil utilisé, le LI-6400 (LICOR®, USA), mesure les échanges gazeux d’une feuille insérée dans une chambre dont les paramètres tels que la lumière, le taux de CO2, l’humidité et la température sont contrôlés. Ainsi, il calcule la concentration de CO2 interne (Ci), la photosynthèse(A) et la conductance de la feuille. Le Vcmax et le Jmax sont estimés par
Figure 1: sources de champs magnétiques (gauche) et pointeur (droite). ^ 200 400 600 800 1000 1200 05 1 0 1 5 2 0 Ci A
Schéma 2: Ajustement des données de photosynthèse d’un hêtre de la parcelle 1 selon les paramètres de Bernachi et al.
Pour cette feuille, Vcmax=46.3167 µmol.m-².s-1 et
9 ajustement des courbes de l’assimilation(A) ou photosynthèse par rapport au CO2 interne(Ci). La lumière est fixée à 1200 µmol.m-².s-1 soit le taux de lumière correspondant à un optimum de photosynthèse et une température de 25 degrés soit la température pour laquelle la Rubisco a un fonctionnement optimal. Les données sont ensuite ajustées à l’aide du logiciel de statistiques R et suivant les paramètres de Bernachi et al. 2001 afin d’obtenir deux courbes de régression (cf. schéma 2): la première s’alignant sur les points proches de l’origine et la seconde s’alignant sur le plateau de points. La pente de la première courbe indique le Vcmax à la température de 25 degrés et celle de la seconde courbe indique le Jmax à 25 degrés. La respiration à l’obscurité(Rd) peut être estimée comme étant la valeur de X pour Y=0 de la courbe A/Ci.
Chaque feuille mesurée est ensuite collectée et séchée à l’étuve à 80°C pendant 60 heures puis réduite en poudre fine au broyeur à bille. Auparavant, sa surface foliaire est mesurée au planimètre. 3 mg de poudre sont prélevés et passés au microanalyseur. La sublimation de la matière sèche permet de détecter le carbone sous forme gazeuse de CO2 et l’azote en N2. Le résultat obtenu est exprimé en masse d’azote par unité de surface foliaire(Na).
b. Mesures de l’interception de la lumière
Une digitalisation 3D des plants de hêtres permet de calculer leur efficience d’interception de la lumière.
Chaque feuille est détectée à travers un champ magnétique émis par une source placée à proximité de l’arbre. Un pointeur est placé à la jonction limbe-pétiole de chaque feuille en le tenant parallèle au plan du limbe (cf. Figure 1). La tête du pointeur est une pointe en plastique sur laquelle sont fixées 3 bobines électromagnétiques dont le courant induit dépend du champ magnétique de la source. Ainsi, la position et l’orientation de chaque feuille en 3D sont enregistrées dans un tableur de type Excel du logiciel PiafDigit. La longueur de chaque feuille(L) est également mesurée et
10 ajoutée au tableur en vue du calcul des LA. Ces données sont ensuite traitées via le logiciel Vegestar afin de calculer le STAR selon une séquence nommée « Turtle 46W ». Le STAR correspond au rapport entre la surface foliaire projetée orthogonalement à la direction de la source de la lumière et la surface foliaire totale, soit, la fraction de la surface foliaire qui intercepte la lumière dans une direction donnée. C'est l'efficience d'interception de la lumière. La séquence « Turtle 46 » permet de calculer le STAR pour 46 directions différentes autour du plant qui représentent les 46 fractions de l’hémisphère du ciel en secteurs d’angles solides égaux. La surface de chaque feuille est calculée à partir de sa longueur, la largeur(l) étant considérée comme proportionnelle à la longueur selon la relation L=k.l. Le coefficient k est estimé, selon la bibliographie, comme étant égal à 0.7 pour le hêtre. Il a été vérifié par les mesures de longueur et largeur de plus d’une trentaine de feuilles par arbre digitalisé et corrigé (il varie entre 0,66 et 0,73 au lieu de 0,7) pour le traitement des données par Vegestar. La longueur et la largeur des feuilles sont mesurées suivant et perpendiculairement à la nervure principale. Le nombre de feuilles et la surface foliaire ou LA sont calculés automatiquement par Vegestar. L’éclairement transmis au-dessus des hêtres a été mesuré, au préalable, par des capteurs de lumière sur une période de 24 heures et rapporté à un témoin en pleine lumière afin de s’affranchir des variations météorologiques qu’il peut y avoir sur une période de 24 heures.
c. Mesures dendrométriques
Le diamètre de la tige principale à quelques centimètres du sol est estimé au pied à coulisse pour chaque hêtre par deux mesures perpendiculaires l’une à l’autre du fait de sa forme ovoïdale. La hauteur est mesurée à l’aide d’une perche depuis la base du tronc jusqu’à la cime.
Figure 2: relations azote foliaire surfacique/éclairement et Vcmax, Jmax/azote surfacique.
Une augmentation de la concentration en azote par unité de surface de feuille en fonction de PARt est observée de même qu’une relation positive entre les deux paramètres de photosynthèse Vcmax et Jmax avec la concentration d’azote par unité de surface de feuille. Les courbes représentées correspondent aux années 2001( ), 2002( ), 2003( ), 2004( ), 2006( ) et 2008( ). 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 0 2 4 6 8 10 accroissement de diamètre en mm STA R
Figure 3: Evolution du STAR en fonction de l’accroissement de diamètre.
On observe une diminution du STAR avec l’augmentation du diamètre de la tige principale. Ce phénomène semble s’accentuer avec les années. Les courbes représentées correspondent aux années 2001( ), 2002( ), 2003( ), 2004( ), 2006( ) et 2008( ).
Figure 4: Accroissement de diamètre et quantité d’éclairement réellement interceptée.
La pente de chaque courbe représente le coefficient d’efficience d’utilisation de la lumière ε pour chaque année de croissance. Il a diminué de 2002 à 2008.
Résultats
Na, Vcmax et Jmax augmentent avec la disponibilité en lumière conditionnée par le couvert forestier (PAR) (cf. Figures 2 et 5). Une relation positive entre Vcmax, Jmax et Na est observée. En comparaison avec les précédentes années, Na, Vcmax et Jmax sont plus élevés.
L'accroissement du diamètre de la tige principale entraîne une diminution du STAR ou efficience de l'interception de la lumière (cf. Figure 3). Ce résultat est observé au long des 7 années d’expérimentation (2001-2008) mais s’amplifie pour les dernières années. Le produit PARt*STAR*LA est l’interception de la lumière par la surface foliaire totale et exacte du plant(LA) modulée par la surface réellement interceptée (STAR) et l’éclairement du milieu(PARt), c’est la quantité d’éclairement réellement interceptée par la plante. Une augmentation de ce produit augmente l'accroissement du diamètre (cf. Figure 4).
Le nombre de feuilles et, par conséquent la surface foliaire totale du plant, augmente avec le PARt. L'angle d'inclinaison moyen des feuilles tend vers l'horizontale à faible éclairement moyen du plant (PARt*STAR) et est plus verticale à fort éclairement (cf. Figure 6). Enfin, la masse de matière sèche par unité de surface foliaire(LMA) augmente avec l’éclairement (cf. Figure 7).
Le coefficient ε, pente de la courbe d’accroissement de diamètre/PARt*STAR*LA, est le coefficient d’efficience d’utilisation de la lumière. Il a diminué entre 2002 et 2008 (2.34 à 0.74) (cf. Figure 4).
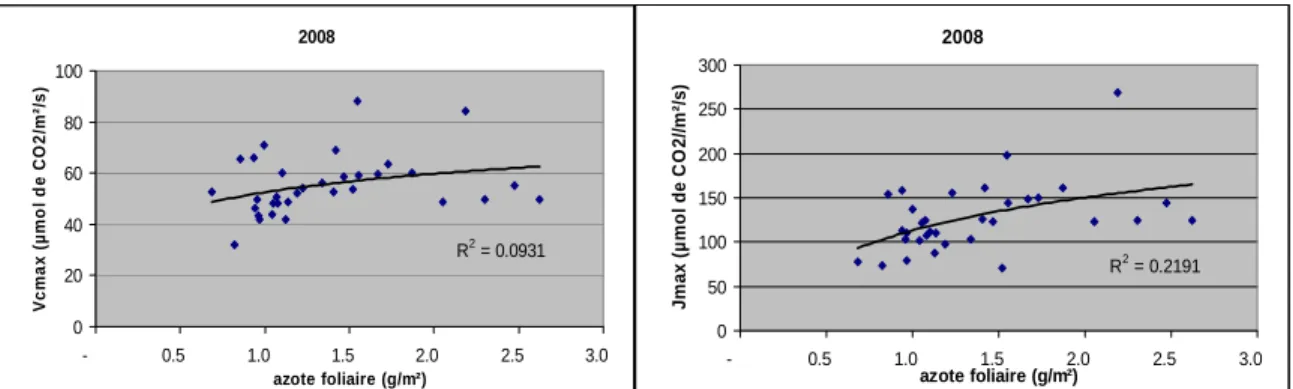
2008 R2 = 0.0931 0 20 40 60 80 100 - 0.5 1.0 1.5 2.0 2.5 3.0 azote foliaire (g/m²) V c ma x (µmo l d e CO 2 /m²/ s ) 2008 R2 = 0.2191 0 50 100 150 200 250 300 - 0.5 1.0 1.5 2.0 2.5 3.0 azote foliaire (g/m²) Jm ax ( µ m o l d e C O 2/ /m ²/ s)
Figure 5: Evolution des deux paramètres de photosynthèse Vcmax et Jmax en fonction de la concentration en azote foliaire surfacique.
Ces deux paramètres augmentent avec la quantité d’azote foliaire.
Figure 6: Variations de l’angle d’inclinaison en fonction de l’éclairement moyen.
L’angle d’inclinaison des feuilles augmente avec l’intensité de l’éclairement du plant. Il tend vers la verticale à fort éclairement et vers l’horizontale dans le cas inverse.
R2 = 0.8129 0 10 20 30 40 50 60 70 80 0 0.2 0.4 0.6 0.8 1 PARt m a tièr e sèche fo liair e (g/m ²) 2008 Logarithmi que (2008)
Figure 7: Masse de matière sèche par unité de surface foliaire en fonction de l’éclairement.
12
Discussion
L'azote est principalement investi dans les protéines, or la Rubisco qui est l'enzyme clé de la photosynthèse représente plus de 50% de ces protéines. Une quantité élevée d'azote par unité de surface foliaire signifie donc une quantité élevée de Rubisco et, par conséquent, une efficacité d'utilisation de la lumière élevée. L'azote augmentant avec le PARt, Vcmax et Jmax étant liés à la concentration en azote surfacique, il peut être considéré que ces deux derniers paramètres sont liés au PARt. L'analyse de ces données sur plusieurs années montre une augmentation à la fois en azote surfacique et en Vcmax et Jmax. Les capacités photosynthétiques augmentent donc avec la lumière et d’année en année.
L’analyse de la respiration foliaire n’est pas assez précise, l’estimation restant difficile, pour être interprétée. En effet, l’estimation par ajustement de droites n’est pas assez précise et les conditions de mesures in situ ne sont pas assez rigoureuses pou être prises en compte.
Le nombre de feuilles, la surface totale foliaire, la variation des angles d'inclinaison des feuilles et l’arrangement spatial et global du feuillage, plane et dispersé en milieu ombragé et tendant plus vers la verticale et plus dense en milieu très éclairé sont des paramètres influant sur le STAR et dépendent de l'éclairement disponible conditionné par le couvert forestier (PARt), mais aussi de la dimension des individus, les feuilles étant plus superposées chez les grands individus.
Le jeune plant répond aux conditions d'éclairement du milieu en conservant un bilan de carbone positif. En milieu sombre, la lumière disponible étant faible (PARt proche de 0), l'efficience de son interception est nécessairement élevée. En milieu très éclairé (PARt proche de
13 1), la lumière disponible étant importante, l'efficience d'interception est faible afin de diminuer les risques de photoinhibition.
De plus, le coefficient d’efficience d’utilisation de la lumière ε a diminué d’année en année. Les résultats de 2001 ne sont pas pris en compte puisqu’il s’agit de l’année d’implantation des jeunes plants de hêtres tous issus de pépinière et l’année 2003 a été une année où la croissance des hêtres a été limitée par de fortes conditions de sécheresse, le coefficient ε n’a donc pas de signification pour 2003. .
Il indique que plus l’arbre grandit et plus l’encombrement spatial est important car la densité du feuillage et donc le recouvrement des feuilles augmentent. En conséquence, le STAR diminue. L’efficience de d’interception de la lumière diminue.
Les tissus non-photosynthétiques augmentent plus rapidement que les tissus photosynthétiques, d’où une diminution du LAR. Il faut donc de plus en plus de carbone pour entretenir la respiration de l’arbre. Enfin, il peut en être déduit que si PARt n’augmente pas, ε diminue.
La plasticité des traits physiologiques et morphologiques du hêtre lui donne son caractère de tolérance à l'ombrage mais celui-ci s’atténue avec l’augmentation de la dimension des individus et donc la diminution de l’efficacité d’utilisation de la lumière. La tolérance d’un arbre à l’ombrage serait donc de limiter la croissance et donc les dimensions (dimension critique) pour que le bilan de carbone reste toujours positif.
14
Conclusion
La tolérance à l’ombrage des jeunes hêtres est un atout pour leur régénération mais leur croissance exige une disponibilité en lumière progressive. La tolérance à l’ombrage n’est plus définie comme la capacité à croître sous un faible éclairement mais comme la capacité de survivre en ralentissant la croissance et reprendre une activité normale après découvert. Les capacités photosynthétiques s’améliorent avec la croissance mais l’augmentation des dimensions globales du plant nécessite, en théorie, une plasticité des traits morphologiques pour optimiser l’efficacité d’interception de la lumière. En pratique, la réponse du hêtre à l’ombrage est une diminution de la croissance.
Une analyse plus approfondie sur les traits morphologiques (les parties ligneuses) permettrait de mieux comprendre le développement des jeunes plants de hêtres et savoir si la notion de tolérance à l’ombrage comprend l’orientation de croissance des tiges secondaires vers des points de lumière (tels des trouées dans la canopée) ou si elle ne concerne que les traits morphologiques foliaires vus dans les précédentes études.
15
Références bibliographiques
Balandier, P., C. Collet, et al. (2006). "Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation." Forestry
79(1): 3-27.
Balandier, P., H. Sinoquet, et al. (2007). "Six-year time course of light-use efficiency, carbon gain and growth of beech saplings (Fagus sylvatica) planted under a Scots pine (Pinus sylvestris) shelterwood." Tree Physiol 27(8): 1073-82.
Bernacchi, C.J., E.I. Singsaas, C. Pimentel, A.R.J. Portis & S.P. Long 2001. Improved temperature response functions for models of Rubisco limited photosynthesis. Plant, Cell and Environment. 24: 253-259.
Farquhar, G. D., S. Von Caemmerer, et al. (1980). "A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species." Planta 149: 78-90.
Fleck, S., U. Niinemets, et al. (2003). "Three-dimensional lamina architecture alters light-harvesting efficiency in Fagus: a leaf-scale analysis." Tree Physiology 23(9): 577-589.
Harley, P. C. and J. D. Tenhunen (1991). Modeling the photosynthetic response of C3 leaves to environmental factors. Modeling crop photosynthesis-from Biochemistry to Canopy. A. S. o. A. a. C. S. S. o. America. Madison, USA. CSSA 19: 17-39.
16 Prevosto, B. and P. Balandier (2007). "Influence of nurse birch and Scots pine seedlings on early aerial development of European beech seedlings in an open-field plantation of Central France." Forestry 80(3): 253-264.
17
Annexes
Année Coefficient ε R² 2001 0.92 0.47 2002 2.34 0.67 2003 0.61 0.32 2004 1.17 0.65 2006 1.28 0.75 2008 0.74 0.55 Annexe 1: Coefficients ε de 2001 à 2008. 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 1 2 3 4 5 6 7 8 9 10 11 12 valeur ordonnée Lu m iè re Série1Placette Hêtre Lumière
1 48 0.066 1 55 0.023 2 37 0.094 2 72 0.138 3 19 0.17 3 33 0.288 4 14 0.259 4 20 0.29 5 5 0.179 5 86 0.121 6 84 0.8 7 43 0.9
18 Ce stage m’a fait découvrir la recherche en milieu forestier. Travailler sur le terrain a été une nouvelle et enrichissante expérience autant sur le plan scientifique que humain qui complète ma formation universitaire en joignant la pratique à la théorie. J’ai ainsi pu approfondir mes connaissances en écologie.
C’est pourquoi je tiens à remercier Monsieur Philippe Balandier pour m’avoir donné l’occasion de réaliser ce stage, ainsi que pour ses conseils et nombreuses explications. Je
remercie également Marc Vandamme et André Marquier qui m’ont guidée tout au long du stage pour m’apprendre à réaliser des mesures de photosynthèse et de digitalisation et de traiter les données.