The Bimodular G57-V577 Polypeptide Chain of the Class B Penicillin-Binding Protein 3 of Escherichia Coli Catalyzes Peptide Bond Formation from Thiolesters and Does Not Catalyze Glycan Chain Polymerization from the Lipid II Intermediate
Texte intégral
Figure
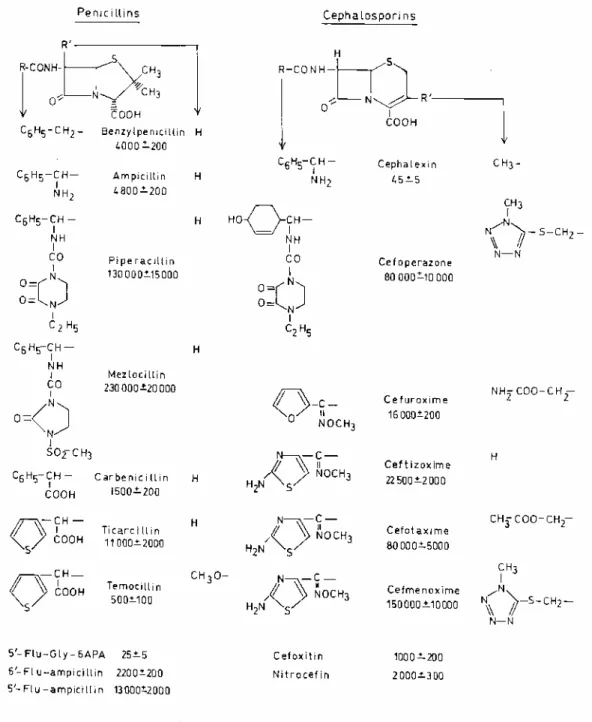
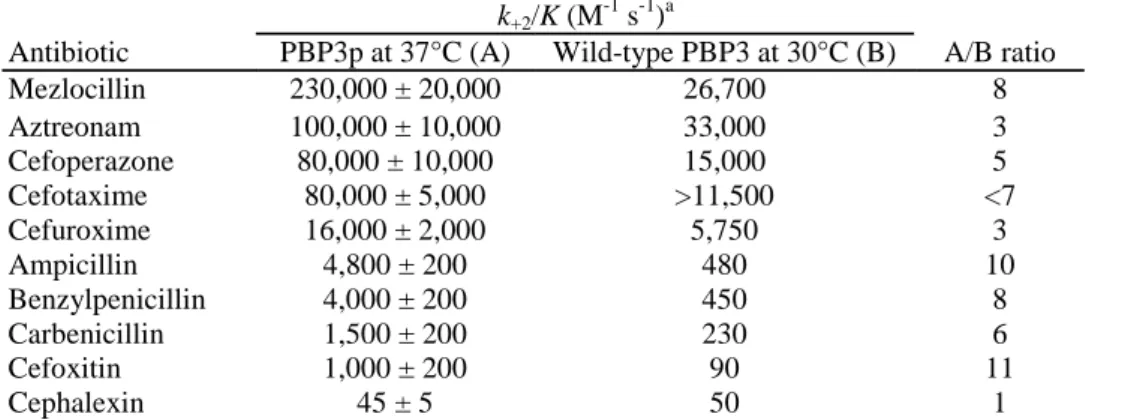
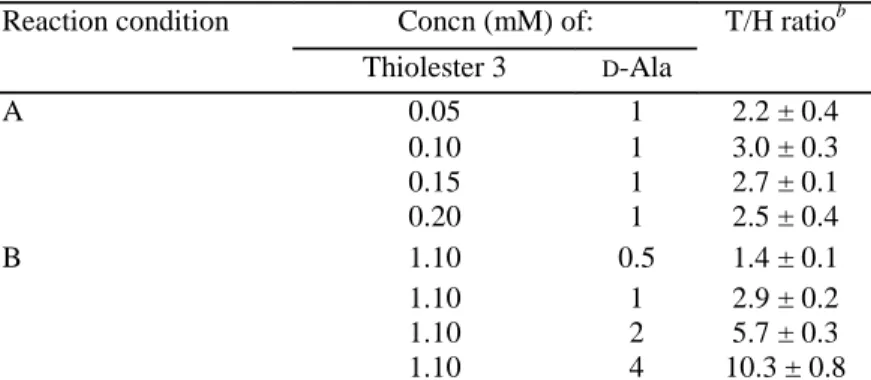
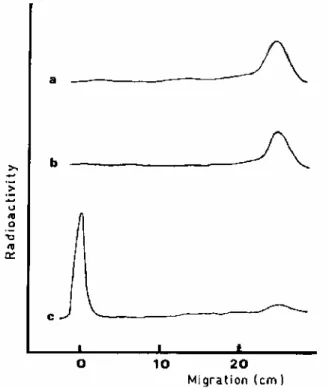
Documents relatifs
Interestingly, when dehydrated sibutramine hydrochloride was recrystallised in anhydrous methanol, and rehydrated in ambient conditions, a single melting peak was also observed by
In contrast to WT female mice, for which we identi fied several PPAR a-associated genes as up-regulated in mice fed pesticide chow compared to those fed control chow ( Figure 5E ),
Ainsi, si la demande globale de bien « énergie » E constitue une contrainte pour ces dernières, la répartition de E entre bien dangereux et bien non dangereux est
We will focus on the in fluence of the aspect ratio of NN NW’s and NN P’s piezoelectric particles as well as the dielec- tric permittivity of the matrix on the piezoelectric
Stochastic partial differential equations; Nonlinear Schrödinger equation; White noise dispersion; Numerical methods; Geometric Numerical Integration; Exponential
- قفتت بلغأ تافيرعتلا ىلع نأ ةيافكلا ةردق ةبستكم - دجوي لخادت نيب ةيافكلا ةراهملاو ءادلأاو فدهلاو ،يكولسلا ثيحب بعصي زييمتلا ،اهنيب دكؤيو كلذ ام بهذ هيلإ
[r]
Hybrid regimes should be understood as possessing various combinations of these democratic and autocratic tendencies, as well as their own distinct attributes.
