Génétique évolutive
Qu'est-ce que la génétique des populations?lois de la génétique de la transmission des caractères ➢caractères à déterminisme simple
théorie de l'évolution (évolution des espèces , sélection naturelle)
➢caractères à
Qu'est-ce que la génétiquedes populations?
Information génétique portée par des locus présentant plusieurs états alléliques
distincts
Polymorphisme floral chez les Gueules-de-loup (Antirrhinum majus)
groupes géographiques d'individus susceptibles de se reproduire entre eux
unité évolutive de l'espèce
Exemples:
•Tous les éléphants d'un parc national donné •Tous les chênes d'un massif forestier donné
(pollen à large dispersion anémogame) •Tous les individus d'une espèce de parasite intestinal, présents chez un seul individu hôte
Variation génétique s'exprime par les fréquences relatives des différents allèles
Evolution se traduit par une variation des fréquences alléliques dans les populations Forces évolutives = processus qui agissent sur les fréquences alléliques
Objectifs:
(1) Mesurer variation génétique dans populations naturelles + décrire patron d'organisation de variation
(2) Expliquer origine, maintien et évolution de la variation génétique par effet des forces évolutives
La génétique des populations est basée sur l’existence d’une variabilité génétique elle concerne de nombreux gènes avec plusieurs allèles (Ex. couleur des yeux, isoenzymes, mutations ponctuelles…), ou des changements de fréquences alléliques (évolution des populations, des espèces)
Mesure de la variabilité génétique dans et entre les populations
– Retrace l’histoire évolutive des espèces , Quantification – description
Explique les modifications observées dans le temps et l’espace : forces évolutives Explication Prédiction
Mesure de la variabilité génétique dans et entre les populations
– Retrace l’histoire évolutive des espèces , Quantification – description
Explique les modifications observées dans le temps et l’espace : forces évolutives Explication Prédiction
Pourquoi s’intéresser au niveau « population » ?
❑Une unité écologique ❑ Une unité évolutive
❑ Individus en interaction avec l’environnement La génétique des populations est probabiliste : ❑ Grand nombre d’individus
❑ Grand nombre de générations ❑ Grand nombre de populations ❑ Facteurs évolutifs multiples Difficile d’expérimenter !
Elle nécessite donc souvent l’utilisation de simulations et modélisations (et de quelques calculs de probabilités)
La théorie de l’échantillonnage :
Echantillon – estimation de la fréquence des allèles Extrapolation à la population globale
« Rien en biologie n’a de sens, … si ce n’est à la lumière de l’évolution » Th. Dobzhansk En Systématique (classification des êtres vivants)
En physiologique (fonctionnement des êtres vivants) - Explique la mise en place des fonctions (hasard et nécessité)
En biologie moléculaire (évolution moléculaire) etc…
La génétique évolutive recherche la cause ultime d’un phénomène (par opposition aux causes proximales), elle répond aux questions : Pourquoi cet allèle et quel est son
devenir.
La notion d'espèce est d'autant plus importante en biologie qu'elle constitue l'unité taxinomique fondamentale évolutive la plus évidente pour tous.
Son utilisation dans plusieurs domaines de la biologie en précise l'intérêt : -Les taxinomies et les systématiques reposent souvent sur la notion d'espèce.
-Le raisonnement génétique est fondé sur la transmission d'allèles chez des individus d'une même espèce et rarement au niveau du genre ou de la famille.
-L'étude de l'évolution se rapporte à celle de l'espèce.
-L'écologie utilise abondamment la notion d'espèce : reconstitution de réseaux trophiques, prévisions des conséquences de pollutions, lutte biologique ou non contre des ravageurs... - La recherche médicale, en parasitologie par exemple, repose sur une définition très précise de l'espèce. C'est ainsi que la recherche d'un vaccin antimalaria a permis de reconnaître six espèces jumelles, chez le Moustique vecteur (Anopheles maculipennis). Elle a pu alors
Ce chapitre étudie la spéciation, c'est-à-dire les processus qui conduisent à l'apparition de nouvelles espèces. Mais, avant d'envisager les modèles et les mécanismes de la spéciation, il est nécessaire tout d'abord de préciser quels sont les critères utilisés pour définir l'espèce.
Les critères spécifiques et leur discussion
La notion d'espèce repose sur plusieurs critères : morphologique,
biologique, écologique
Définition de l’espèce biologique :
L'espèce biologique est constituée «
de groupes de
populations
naturelles
se
croisant
librement
ou
potentiellement capables de le faire, et isolées
reproductivement
des groupes appartenant aux espèces
voisines ».
Remarquons que le critère morphologique est totalement
exclu de cette définition.
LES MECANISMES D'ISOLEMENT Définition et caractérisation
Les espèces voisines se différencient les unes des autres par toute une série de caractères morphologiques, physiologiques et génétiques.
Au niveau de chaque individu d'une espèce donnée, il existe des mécanismes qui suppriment, ou ne permettent normalement que très modérèrent, les échanges génétiques entre les différent membres d'espèces différentes.
On donne le nom de mécanisme d'isolement a de telles barrières, dont la mise en œuvre a pour résultat de maintenir l'intégrité spécifique.
Différentes catégories de mécanismes d'isolement L'isolement géographique
c'est-a-dire la simple séparation dans l'espace, aboutit également a une suppression des échanges entre espèces,
Une nouvelle espèce naît après qu'une population se soit retrouvée isolée sur le plan géographique de sa population ancestrale.
Il y a fragmentation de l'aire de distribution.
La transformation des milieux est le principal responsable de la fragmentation des aires de distribution des espèces.
Dans notre petit schéma théorique qui montre les allers/retours entre le centre de dispersion d'une espèce et les nouveaux milieux colonisés, nous comprendrons
facilement que l'installation d'une barrière autour du centre de dispersion va inhiber le retour des individus migrants et partant le brassage génétique des adaptations acquises dans des milieux différents. Les espèces originellement brassées continuent d'évoluer et de s'adapter localement sans ne plus rien s'échanger.
L'isolement géographique doit être distingue des mécanismes d'isolement proprement biologiques survenant normalement entre les membres d'espèces différentes occupant la même aire. Pour chaque espèce ils sont nombreux et varies;
Dobzhansky, qui a été l'un des premiers auteurs a avoir attire l'attention sur l'importance de ces mécanismes et qui a étudie un certain nombre d'entre eux en détail, a propose de les classer en deux grandes catégories:
les mécanismes pré-zygotiques qui empêchent la formation des zygotes hydrides, et les mécanismes postzygotiques qui empêchent la croissance des hybrides ou provoquent leur stérilité, ainsi que la viabilité et la stérilité de
I. Mécanismes pré-zygotiques :
1. Isolement par l'habitat. 2. Isolement saisonnier. 3. Isolement éthologique. 4. Isolement mécanique. 5. Isolement gamétique. II. Mécanismes post-zygotiques
1. A. Inviabilité des hybrides de première génération. 2. B. Stérilité des hybrides de première génération.
3. C. Inviabilité et stérilité des hybrides de deuxième génération.
Description des principaux mécanismes d'isolement reproductif 1.- ISOLEMENT PAR L'HABITAT
L'isolement par l'habitat, ou isolement écologique, est
fréquent chez les végétaux; il réalise la transition avec
l'isolement géographique et provient du fait que les
individus appartenant a deux espèces différentes ont des
préférences écologiques distinctes.
Un bon exemple est celui de deux espèces voisines de
violette, Viola arvensis et V. tricolor, qui peuvent donner
expérimentalement des hybrides fertiles, mais demeurent
toutefois distinctes dans la nature a cause de leurs
préférences écologiques, v. arvensis étant calcicole et V. tricolore silicole.
Chez les animaux également ce mode d'isolement peut présenter
une certaine importance parce que ceux-ci sélectionnent
parfois les habitats dans lesquels ils vivent et se
ISOLEMENT SAISONNIER
On parle d'isolement saisonnier lorsque les périodes de reproduction de deux espèces ne coïncident pas au cours de l'année. L'exemple classique est celui de deux espèces
voisines de laitue, Lactuc acanadensis et L. graminijolia, qui sont capables de produire des hybrides artificiels
viables et fertiles mais ne fleurissent pas a la même époque de l'année, L. graminiiolia ayant une floraison printanière précoce et L. canadensis fleurissant en été.
ISOLEMENT ETHOLOGIQUE
L'isolement éthologique, dont la catégorie la plus importante est l'isolement sexuel, est très répandu chez les animaux à psychisme développe, la barrière principale a I'accouplernent provenant chez eux d'incompatibilités se manifestant lors du
comportement sexuel; il est fonde sur l'interaction, aux
niveaux de l'émission et de la réception, entre stimulus
provenant des partenaires des deux sexes, ces stimulus étant visuels, auditifs, olfactifs ou tactiles. Par exemple deux espèces de grenouille Microhyla olivacea et M. carolinensis occupent respectivement l'ouest et l'est du sud des Etats-Unis; il a pu être démontré que dans la zone de superposition des deux espèces leurs chants de cour étaient très distincts, et qu'il n'existait pratiquement pas de mécanismes autres que
la voix permettant la séparation des deux groupes. Chez
plusieurs espèces de drosophiles, de blattes, d'autres espèces
d'Insectes et certains Mammiferes, des signaux chimiques
agissant a distance ou par contact peuvent être a l'origine de
mécanismes d'isolement, chaque signal etant plus ou moins
ISOLEMENT MECANIQUE
Chez certaines plantes a f1eur la structure de la corolle ne permet le passage que de catégories spéciales d'insectes
pollinisateurs, réduisant les risques d'hybridation et réalisant ainsi un isolement mécanique d'une notable efficacité, Chez bon nombre d'espèces d'orchidées, la
pollinisation s'effectue par L'intermédiaire d'insectes; la ressemblance de la fleur avec un insecte femelle dans
certains cas entraine l'attraction des mâles qui réalisent ainsi une pseudo-copulation, libérant du pollen sur le
stigmate et recevant en même temps les pollinies. Diverses espèces d'Ophrys sont pollinisées par des mâles d'Andrena et de Scolia, les f1eurs des différentes espèces d'orchidées n‘étant attractives que pour une seule espèce d'insecte. Chez certaines plantes a f1eur la fécondation peut échouer parce que le pollen d'une espèce a styles courts ne peut développer un tube pollinique suffisamment long dans les espèces a longs styles, ou parce que cette croissance y est ralentie ou empêchée.
ISOLEMENT GAMETIQUE
Un certain nombre de cas d'isolement gamétique sont connus; ils consistent en une inévitabilité des gamètes d'une espèce au contact des voies sexuelles ou du style d'une autre. Chez de
nombreuses drosophiles les spermatozoïdes d'une espèce
étrangère provoquent dans le tractus génital de la femelle
réceptrice une réaction antigénique; cette réaction a
l'insémination aboutit a un gonf1ement du vagin et a la mort des spermatozoïdes.
Apres la fécondation, les trois barrières principales sont
l'inviabilité des hybrides de première génération, la stérilité de ces hybrides lorsqu'ils sont parvenus a l'etat adulte, et
l'inviabilité ou la stérilité des hybrides des générations
ultérieures. Selon Stebbins, cette dernière pourrait être « de développement » -les gonades se développant anormalement, ou la méiose ne pouvant s'effectuer - ou « de ségrégation » - la
ségrégation de combinaisons de gènes, de segments
chromosomiques, ou de chromosomes entiers dans les gamètes ne se réalisant pas de façon correcte.
INVIABILITE DES HYBRIDES DE PREMIERE GENERATION
Une des études les plus fouillées relative a I‘inviabilité des hybrides de première génération est celle que Moore a effectué sur l'hybridation entre douze espèces de
grenouilles d'Amérique du nord appartenant au genre Rana; l'arret de développement des hybrides survient selon les croisements a différentes phases critiques au cours de la période embryonnaire : avant les premières divisions de l’œuf fécondé, a la gastrulation, ou a un stade ultérieur.
STERILITE DES HYBRIDES DE PREMIERE GENERATION
L'exemple le plus célèbre de stérilité des hybrides de première génération est celui du mulet qui, bien qu'etant un animal
particulièrement résistant, n'en est pas moins complètement
stérile; cette stérilité des hybrides de première génération est un phénomène très général. Même lorsque les hybrides de premiere génération entre deux espèces sont viables et féconds, il peut exister une barrière aux échanges de matériel génétique lors des générations ultérieures; par exemple, bien que les hybrides de première génération entre les diverses espèces de cotons du genre Gossypium (tels que G. hirsutum, G. barbadense et G.
tomentosum) soient vigoureux et fertiles, une très forte proportion des semences de seconde génération ne germe pas.
Les espèces ont donc generalement a leur disposition tout un eventail de possibilites autres que le seul precede de sterilite pour conserver leur identite. Pour une espece donnee, differents mecanisrnes se completent et s'epaulent; deux especes de
drosophiles par exemple seront separees par une combinaison d'isolement ethologique, d'inviabilite hybride partieIle et de sterilite hybride chez le male;
deux especes de plantes seront quant a elles separees par des preferences ecologiques cornbinees a des differences dans la structure de la corolIe, une inviabilite et une sterilite des hybrides. Certains mecanismes sont composites, comme J'isolement ethologique qui peut etre subdivise en plusieurs sous-categories. Bien sou vent I'un des mecanisrnes predomine sur les autres (il en est ainsi en particulier pour l'inviabilite et la sterilite des hybrides de premiere generation), et cette propriete revet des modalites variables selon les especes,
6. LA SPECIATION
6.1. Classification des modalités de spéciation.
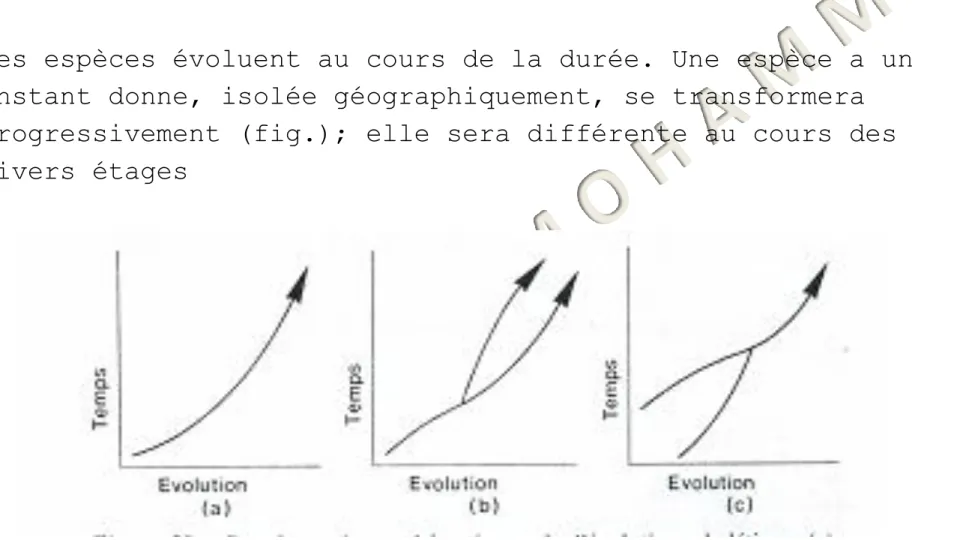
Les espèces évoluent au cours de la durée. Une espèce a un instant donne, isolée géographiquement, se transformera progressivement (fig.); elle sera différente au cours des divers étages
Figure Représentations schématiques de l‘évolution phylétique (a), la spéciation primaire (b), et la spéciation secondaire (c)
du temps géologique, la désignation précise du niveau de la séparation entre les « espèces » successives étant bien entendu arbitraire.
On donne le nom d‘évolution phylétique a ce mode de
spéciation, et il ne s'accompagne pas a un instant donne d'une augmentation du nombre des espèces.
Le terme de spéciation concerne en effet habituellement les modalités aboutissant a une multiplication du nombre des espèces,
Les termes d'anagenèse et de cladogenèse concernent
respectivement l‘évolution phylétique opposée a la division d'une espèce en deux ou plusieurs espèces nouvelles; il est possible dans ce dernier cas de distinguer deux modalités :
soit il y a division complète, soit une ou plusieurs
Deux especes peuvent fusionner completement.
Cette fusion se realise par rupture des mecanismes
d'isolement qui protegent normalement les especes.
Ce processus est designe sous le vocable de speciation
secondaire, car il s'accompagne d'une reduction du
nombre des especes et necessite l'existence prealable
d'au moins deux d'entre elles. A l'interieur de la
speciation par multiplication, Mayr a propose de
distinguer la speciation instantanee de la speciation graduelle (tableau).
La speciation instantanee, ou individuelIe, est definie comme la production des descendants d'un seul couple,
reproductivement isoles de l'espece a laquelle
appartient la souche parente, et capables d'etablir une nouvelle population donnant ulterieurernent naissance a une espece;
les sous-categories de la speciation instantanee concernent les modalites utilisees pour aboutir a une telle production,
modalites qui sont au nombre de deux principales : la transformation par restructuration chromosomique et la
transformation par polyploidie. La speciation graduelle est la divergence progressive des populations jusqu'à l'atteinte du niveau permettant la distinction specifique; il existe deux grandes sous-categories de speciation graduelle : la speciation allopatrique faisant intervenir l'isolement geographique, et la speciation sympatrique, sans isolement geographique. En fait, hybridation, restructuration chromosomique et polyploidie sont egalernent considerees au niveau des populations lorsque se pose le problerne des transformations genetiques en cours de
speciation, il est probable qu'a l'echelle du temps geologique, le dessin complexe de disparition et d'apparition d'especes
voisines se presente sous la forme d'une intrication reticulaire compliquee, flux genique, introgression, hybridation, fusion et amphiplorde constituant autant de procedes tissant la partie transversale des mailles du reseau
Tableau : Classification des diverses modalites de speciation (d'apres MAYR). I. Evolution phylétique. 2. Fusion. 3. Multiplication A. Instantanée :
• par restructuration chromosomique, • par polyploidie.
B. Graduelle :
• sympatrique, • allopatrique.
Généralisation du modèle de Hardy-Weinberg
Sous le modèle de Hardy-Weinberg, la composition génétique d’une population demeure
stable, inchangée d’une génération à l’autre, si
trois conditions sont réunies :
1. absence ou effet négligeable sur quelques générations, des mutations, de la sélection et des migrations ;
2. taille infinie ou assez grande pour que les fréquences des évènements soient égales à leurs probabilités (loi des grands nombres). Sous ces deux conditions, les fréquences alléliques demeurent inchangées ;
3. panmixie (incluant la condition de pangamie), c'est-à-dire croisements au hasard.
Sous cette condition, une relation mathématique, désignée par relation panmictique ou relation de Hardy-Weinberg, permet de calculer les fréquences génotypiques à partir des seules fréquences alléliques.
La dérive génétique
Le modèle de Hardy-Weinberg suppose que l’effectif des populations
est suffisamment grand pour être considéré comme infini.
Cette condition est évidemment irréaliste, mais, avant de définir un
seuil
à
partir
duquel
cette
condition
serait
en
pratique
acceptable, il convient d’établir ce qu’il advient de la composition
génétique des populations quand l’effectif est limité.
On montre que la limitation de l’effectif d’une population conduit à un
phénomène appelé « dérive génétique » parce qu’il est caractérisé par
une fluctuation aléatoire, d’une génération à l’autre, des fréquences
alléliques.
On montre aussi que le même phénomène peut être caractérisé par une
augmentation récurrente de la consanguinité
Quand la population est de grand effectif, le nombre de tirages est tellement élevé que la fréquence des gamètes tirés ne sera pas beaucoup, voire pas du tout différente de leur probabilité de tirage, soit p et q.
Au contraire, dès que l’effectif sera assez petit, les fréquences des allèles A1 et A2, après le tirage peuvent différer considérablement de leurs probabilités respectives de tirages.
Dans une petite population, les fréquences alléliques vont donc fluctuer au gré des
variations aléatoires de tirages, et ce, d’une génération à l’autre.
Formulation mathématique de la dérive génétique
Si on considère une population d’effectif constant N, on conçoit qu’il faut réaliser 2N tirages à chaque génération, dans l’urne gamétique des parents.
Si les deux allèles A1 et A2 d’un gène di-allélique, ont pour fréquence pi et qi à la génération
i, quelles seront leurs valeurs à la génération suivante ?
En fait, il suffit de s’intéresser à la fréquence d’un seul allèle, celle de l’autre étant le
complément à 1. La probabilité de tirage de l’allèle A1 dans l’urne gamétique est égale à la fréquence pi de l’allèle A1 chez les parents.
Si on fait 2N tirages dans l’urne gamétique le nombre d’allèles A1 tirés peut être compris entre 0 et 2N. On sait plus précisément que chacune des valeurs de cette variable aléatoire Xi + 1 a une probabilité d’observation donnée par la loi binomiale B(pi , 2N). Cette loi permet de définir les paramètres de la distribution de Xi + 1
– son espérance (appelée aussi moyenne) est égale à : E(Xi + 1) = 2N. Pi
On montre bien ainsi que, dans une grande population idéale (hypothèse du modèle de Hardy-Weinberg), quand N tend vers l’infini, la variance de la fréquence
fi + 1 est nulle ; de ce fait la fréquence fi + 1 de A1 ne peut qu’être égale à son espérance pi,
soit la fréquence de A1 à la génération précédente.
Au contraire, dès que l’effectif est limité, la variance n’est plus nulle, et la fréquence fi + 1 à la génération i + 1, peut prendre une valeur différente de la valeur pi à la génération précédente.
Cette variation de valeur entre générations ne dépend que du hasard d’échantillonnage.
Comme la variation de la fréquence de l’allèle A1,
de génération en génération est totalement imprévisible, elle fluctue de manière aléatoire ou chaotique.
Cette fluctuation allélique peut être illustrée par la figure, où trois populations,
au départ identiques dans leur composition génétique (même fréquence initiale f0), voient leur diversité évoluer de manière chaotique et divergente en raison de la dérive génétique
Les fréquences alléliques des trois populations A, B et C fluctuent de manière totalement aléatoire entre 0 et 1, les deux valeurs extrêmes possibles de la fréquence, à chaque génération.
Mécanismes de la microévolution