Effets de facteurs naturels et de la coupe de
récupération sur la défoliation de la
régénération préétablie pendant une épidémie
de la tordeuse des bourgeons de l’épinette
Mémoire
Anne Cotton-Gagnon
Maîtrise en sciences géographiques
Maître en sciences géographiques (M. Sc. géogr.)
Québec, Canada
Effets de facteurs naturels et de la coupe de
récupération sur la défoliation de la
régénération préétablie pendant une épidémie
de la tordeuse des bourgeons de l’épinette
Mémoire
Anne Cotton-Gagnon
Sous la direction de :
Martin Simard, directeur de recherche
Daniel Kneeshaw, codirecteur de recherche
iii
Résumé
L’épidémie de la tordeuse des bourgeons de l’épinette (TBE) ayant débuté en 2006 au Québec a désormais affecté plus de 7 million d’hectares. Bien que l’abondance et la vigueur de la régénération préétablie soient des facteurs de haute importance concernant la composition et la productivité futures des forêts, très peu d’études ont ciblé la façon dont celles-ci sont affectées par la TBE. La coupe de récupération post-TBE deviendra un enjeu économique majeur en zone boréale et la réduction de son impact passe par une compréhension des processus écologiques affectés par l’épidémie en cours. La présente étude vise à approfondir les connaissances de l’impact d’une épidémie de la TBE et de la coupe de récupération post-TBE sur la défoliation subie par la régénération préétablie de peuplements mixtes de sapin baumier et d’épinette noire. Les objectifs spécifiques sont d’abord d’évaluer l’effet de la hauteur et de l’espèce des tiges de régénération, ainsi que de la composition du peuplement, sur la défoliation subie par la régénération préétablie. Ensuite, d’évaluer l’effet de la coupe de récupération sur cette défoliation. Les résultats démontrent que la hauteur, l’espèce, la composition du peuplement et la coupe de récupération affectent toutes significativement la défoliation subie par la régénération. Les individus plus grands, les tiges de sapin baumier et toutes les tiges se trouvant dans des sapinières subissent davantage de défoliation. La défoliation des tiges d’épinette noire dans les pessières était radicalement plus élevée dans les sites récupérés comparativement aux sites naturels, ce qui pourrait influencer la composition future de ces peuplements. Davantage d’études long-terme sont nécessaires afin de mieux comprendre la dynamique sous la canopée durant une épidémie de la TBE, en particulier dans des peuplements dominés par l’épinette noire, ce qui permettrait une meilleure gestion écosystémique de nos forêts et une diminution leur susceptibilité à la TBE.
iv
Abstract
Since 2006, the area defoliated by the spruce budworm (SBW) in Quebec has doubled almost every year, and it has now affected more than 7 million hectares. Although advance regeneration abundance and vigor are critical factors determining future forest composition and productivity, very few studies have focused on how they are affected by a SBW outbreak. In the years to come, post-SBW salvage logging will become a major economic issue in boreal forests and reducing its impact involves an understanding of the ecological processes affected by the outbreak in progress. This study aims to determine the impact of SBW outbreaks and of post-outbreak salvage logging on the defoliation sustained by advance regeneration in mixed stands of balsam fir and black spruce. The specific objectives were, first, to assess the effect of regeneration height and species (balsam fir or black spruce), as well as canopy composition, on the defoliation of advance regeneration. Second, to evaluate whether the defoliation sustained by advance regeneration is greater in salvage logged sites or in natural stands of different compositions. Results showed that height and species of regeneration, canopy composition and salvage logging all significantly affected defoliation. Taller advance regeneration, balsam fir advance regeneration, and all regenerating stems in fir-dominated stands sustained more defoliation. Defoliation sustained by black spruce regeneration was much greater in post-outbreak harvested stands than in the understory of natural sites. Harvesting stands with an important component of black spruce regeneration could potentially reduce the future abundance of black spruce, the preferred species for harvesting. Long-term studies are needed to better understand the dynamics in the regeneration layer during a SBW outbreak, particularly in spruce-dominated stands, to conduct better ecosystem-based management and to help reduce the overall susceptibility of our forests.
v
Table des matières
Résumé ... iii
Abstract ... iv
Table des matières ... v
Liste des tableaux ... vi
Liste des figures ... vii
Remerciements ... viii
Avant-propos ... ix
Introduction générale ... 1
CHAPITRE 1 – Post-outbreak salvage logging increases defoliation of black spruce advance regeneration in black spruce-dominated stands ... 6
Résumé ... 7 Abstract ... 8 1Introduction ... 9 2Methods ... 12 2.1 Study area ... 12 2.2 Site selection ... 12 2.3 Sampling ... 13 2.4 Statistical analysis ... 14 3Results ... 16
3.1 Stand composition and defoliation history ... 16
3.2 Factors influencing defoliation of advance regeneration in natural sites ... 16
3.3 Effect of salvage harvest on defoliation of advance regeneration ... 16
4Discussion ... 18
4.1 Factors influencing defoliation of regeneration in natural sites ... 18
4.2 Effect of harvest on regeneration defoliation ... 18
4.3 Implications for sustainable forest management ... 20
Acknowledgements ... 21
References cited ... 22
Références citées ... 34
vi
Liste des tableaux
Table 1 Results of the mixed model explaining cumulative defoliation (%) of advance regeneration
in natural sites (n = 2195). N.S. = not significant (alpha = 5%); df = degrees of freedom; CC = canopy composition.. ... 27
Table 2 Results of the mixed model explaining cumulative defoliation (%) of advance regeneration
in natural and harvested sites (n = 7577). N.S. = not significant (alpha = 5%); df = degrees of freedom. In this analysis, canopy composition was only computed at the stand scale.. ... 28
vii
Liste des figures
Figure 1 Relative (left) and absolute (right) stand composition of the canopy (top) and regeneration
(bottom) for the natural (N) and harvested (H) sites. N.A. = not available. ... 29
Figure 2 Cumulative defoliation history in natural sites (circles) versus harvested sites (triangles).
Open symbols indicate superposed trajectories. ... 30
Figure 3 Predicted defoliation of advance regeneration according to height and species of
regeneration for three stand-scale canopy compositions (top; in black spruce-dominated stands, the relative basal area of balsam fir was less than 2%, averaged 42% in mixed stands, and was greater than 72% in balsam fir-dominated stands), and three neighbourhood
compositions (bottom; black spruce-dominated neighbourhoods have a relative basal area of balsam fir that is 10%, that averages 50% for mixed neighbourhoods, and is greater than 90% for balsam fir-dominated neighbourhoods). Stand-scale composition graphs have been calculated based on the mean value of neighbourhood compositions and vice versa.. ... 31
Figure 4 Probability of sustaining severe (≥ 50%) defoliation as a function of stem height. Shaded areas indicate the 95th confidence interval. ... 32
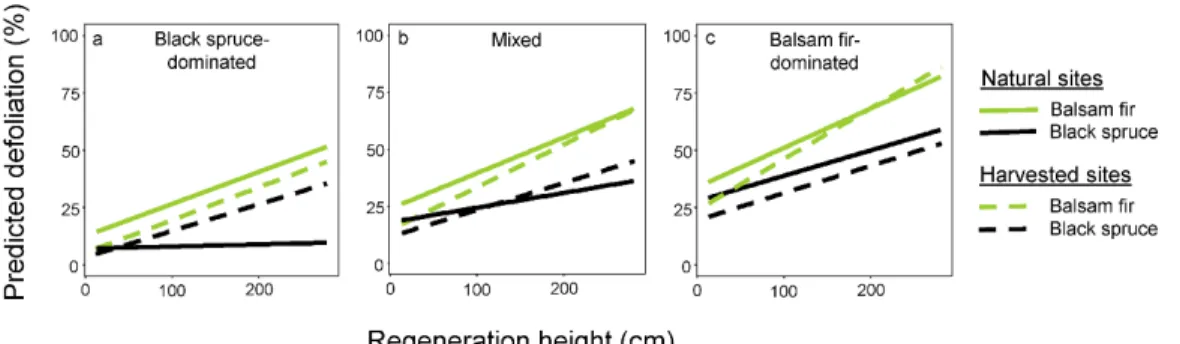
Figure 5 Predicted defoliation of advance regeneration in natural and harvested sites by stem
height for the two studied species and three stand-scale canopy compositions (for black spruce-dominated stands relative basal area in balsam fir is 10%, it is 40% for mixed stands, and 65% for balsam fir-dominated stands).. ... 33
viii
Remerciements
Je voudrais d’abord remercier mon directeur de recherche, Martin Simard, qui m’a trouvé ce merveilleux projet de maîtrise. Martin a été un directeur disponible, dont les nombreux conseils constructifs m’ont permis de continuellement m’améliorer tout en restant confiante et motivée. Je voudrais également remercier mon co-directeur, Daniel Kneeshaw, un homme d’une grande sagesse. Dan est un mentor inspirant qui a su me transmettre sa passion tout en me guidant dans une démarche scientifique efficace. J’aimerais souligner l’apport de Louis De Grandpré, dont la vision et l’expérience ont été les précurseurs de ce projet. Louis fut également très présent, toujours à l’écoute de mes questions, et de bons conseils.
Ce projet étant constitué d’une bonne partie de travaux sur le terrain, il n’aurait certainement pas été possible sans l’aide de nombreuses personnes. Parmi celles-ci figure d’abord Hugues Dorion, qui a coordonné à merveille l’équipe « Jam en Nord » sur la Côte-Nord en 2015 (et plus!) et qui s’est assuré que ma campagne de terrain soit réalisée dans les temps et dans un environnement plus qu’agréable. Également, Gauthier Cervello et Mathieu Gauvin ont été des coéquipiers hors pairs, motivants et motivés, avec qui j’ai passé des moments inoubliables, beau temps, mauvais temps. Un énorme merci à toute l’équipe de terrain qui a participé de près ou de loin à mon projet : Philippe Berthiaume, Stéphane Bourassa, Martín Brasa Estévez, Dave Gervais, Jeanne Moisan-Perrier, Chantale Moulin-Vézina, Roxanne Pommier, Lucas Schmitt, Coralie Serrano et Cédric Valère.
Je me dois également de remercier l’équipe du laboratoire de Christian Hébert, qui m’a soutenue depuis mes débuts en écologie forestière. C’est grâce à mon emploi d’auxiliaire de recherche dans cette équipe que j’ai développé mon intérêt en entomologie forestière. Je voudrais remercier Yves Dubuc et Christian d’avoir partagé leur passion avec moi, ainsi que Jean-Michel Béland, Jonathan Boucher et Olivier Jeffrey d’avoir été de bons conseils et d’un support satisfaisant tout au long de ma maîtrise. J’aimerais aussi remercier Maryse Marchand du laboratoire de Louis De Grandpré pour son aide précieuse en statistiques. Finalement, rien de cela n’aurait été possible ni même envisageable sans les encouragements et le support continu de ma famille. Un énorme merci à mes parents Diane et Conrad, ainsi qu’à mes sœurs Émilie et Marie-Pier, sans qui aucune réalisation ne vaut la peine d’être célébrée. Enfin, le dernier mais non le moindre, merci à mon copain Guillaume d’être mon pilier.
ix
Avant-Propos
Ce mémoire de maîtrise est constitué d’un chapitre, « Post-outbreak salvage logging increases defoliation of black spruce advance regeneration in black spruce-dominated stands ». Ce chapitre rédigé en anglais et sous la forme d’article scientifique est destiné à être publié dans un journal scientifique au cours de l’année 2017. Ce chapitre est précédé d’une introduction rédigée en français. La candidate à la maîtrise a effectué l’installation du dispositif d’échantillonnage du deuxième objectif de l’étude, l’estimation de la défoliation de tous les semis du dispositif, la description des attributs de l’habitat dans les sites d’étude et a fait les différentes analyses statistiques. Le chapitre a été écrit entièrement par la candidate à la maîtrise, encadrée et soutenue par les commentaires et conseils des coauteurs Martin Simard, Daniel Kneeshaw et Louis De Grandpré. L’étudiante a donc le statut d’auteure principale pour l’article paraissant dans ce mémoire et les coauteurs, énumérés à la phrase précédente, ont été impliqués également dans la conception de l’article. Cette étude a été financée par le Fonds de Recherche Nature et Technologies (FRQNT).
1
Introduction générale
Les perturbations naturelles sont généralement acceptées comme étant les moteurs de la dynamique des écosystèmes forestiers terrestres (Pickett 2012). Différents régimes de perturbations entrainent différentes dynamiques dans les populations végétales (Bergeron 1991, Gauthier et al. 1993). Dans la forêt boréale, plusieurs espèces arborescentes ont développé des mécanismes de régénération par reproduction végétative (Zasada et al. 1992), tels que les drageons ou les rejets de souche (Bergeron and Charron 1994, Lavertu et al. 1994), qui leur permettent de persister dans un peuplement et de réenclencher la succession (Holling 1986). Les espèces pionnières telles que le peuplier faux-tremble (Populus tremuloides [Michx.]) ou le bouleau à papier (Betula papyrifera [Marsh.]) (Dix and Swan 1971, Rowe 1983, Johnstone et al. 2004) facilitent l’établissement d’espèces plus tardives telles que l’épinette blanche (Picea glauca [Moench]), l’épinette noire (Picea
mariana [Mill.]), le thuya occidental (Thuja occidentalis [L.]) et finalement, le sapin baumier
(Abies balsamea [Mill.]) (Dix and Swan 1971, Rowe 1983, Kneeshaw and Bergeron 1996, Galipeau et al. 1997, Bergeron and Leduc 1998, Gauthier et al. 2000). À noter que la succession post-perturbation dans la forêt boréale se traduit souvent par une convergence de composition vers des espèces conifériennes (Bergeron and Charron 1994, Bergeron 2000, Gauthier et al. 2010).
Différents facteurs abiotiques peuvent influencer la dynamique de ces écosystèmes et entrainer une divergence dans la trajectoire successionnelle. D’abord, l’intervalle entre deux perturbations telles que les feux de forêt joue un rôle critique sur la composition du peuplement (Rowe 1983, Johnstone and Chapin 2006b). En effet, une augmentation du temps écoulé depuis le dernier feu permet un changement dans la composition de la voûte forestière (Frelich and Reich 1995, Bergeron 2000, Gauthier et al. 2000, Lesieur et al. 2002, Lecomte et al. 2005). La sévérité d’une perturbation peut aussi affecter la composition (Pickett et al. 1987, Johnstone and Kasischke 2005, Lecomte et al. 2005, Johnstone and Chapin 2006a) ainsi que la structure du peuplement affecté (Lecomte and Bergeron 2005), en plus de la densité des espèces s’y établissant (Johnstone and Kasischke 2005, Johnstone and Chapin 2006a). Ensuite, l’interaction entre différentes perturbations se déroulant successivement peut résulter en un changement dans la densité (Kneeshaw and Bergeron 1996) ainsi que la composition (Payette et al. 2000) des semis s’établissant après perturbation. Des perturbations anthropiques, telles que les coupes de récupération, suivant un feu de forêt ou une épidémie de la tordeuse des
2
bourgeons de l’épinette (TBE; Choristoneura fumiferana [Mill.]) peuvent même mener jusqu’à une ouverture des pessières noires à mousse dans la forêt boréale (Dussart and Payette 2002, Payette and Delwaide 2003). Finalement, l’étendue de la perturbation peut aussi influencer la densité des semis (Spring et al. 1974, Galipeau et al. 1997) et le patron de recolonisation (Pickett et al. 1987) du peuplement affecté, particulièrement pour les espèces non adaptées à cette perturbation, puisque celles-ci doivent nécessairement recoloniser à partir des zones non perturbées qui contiennent le réservoir de graines. Étant donné que les changements climatiques risquent d’influencer grandement la fréquence et la sévérité des régimes de perturbations telles que les épidémies d’insectes et les feux de forêt (Bergeron and Flannigan 1995, Weber and Flannigan 1997, Fleming and Candau 1998, Weber and Stocks 1998, Gray 2008, Wotton et al. 2010), il faut considérer ces changements lorsqu’on tente de déterminer la trajectoire successionnelle de la régénération d’un peuplement affecté par de telles perturbations.
La principale perturbation de la forêt boréale en Amérique du Nord est le feu de forêt. En général, le cycle des feux y est court et la structure de la forêt est plutôt équienne (Heinselman 1981, Johnson 1992). Cependant, l’est de l’Amérique du Nord fait exception à cette règle (Gauthier et al. 2001, Cyr et al. 2007, Bouchard et al. 2008). En effet, cette région étant particulièrement humide, l’intervalle entre les feux y est souvent supérieur à la longévité des arbres (Kneeshaw and Gauthier 2003). Ce sont plutôt les épidémies d’insectes et les chablis qui constituent les moteurs de la dynamique de la forêt boréale dans l’est de l’Amérique du Nord (Gauthier et al. 2010). En effet, la superficie de forêts perturbées par la TBE est supérieure à celle affectée par les feux de forêt (Kneeshaw 2001). La TBE est un insecte lépidoptère indigène de l’Amérique du Nord qui cause des dommages par défoliation aux conifères en s’alimentant de leurs aiguilles (Martineau 1985). Plusieurs épidémies ayant eu lieu depuis plus de 200 ans sont répertoriées (Blais 1961, Morin 1998, Bouchard et al. 2006). Ces épidémies durent généralement de 15 à 20 ans à l’échelle régionale, et ont lieu à un intervalle de 30 à 40 ans (Royama 1984, Jardon et al. 2003). En période épidémique, une récurrence de la défoliation du feuillage de l’année peut entrainer de la mortalité chez les arbres hôtes (Maclean and Ostaff 1989). Une étude sur l’alimentation de la TBE démontre une susceptibilité maximale pour le sapin baumier, suivi de l’épinette blanche, de l’épinette rouge, et finalement, de l’épinette noire (Hennigar et al. 2008). En effet, la mortalité observée chez le sapin est plus élevée (10 à 100%) que chez l’épinette noire (0 à 80%; Maclean 1980). La TBE préfère également les vieux individus aux jeunes individus (Bergeron et al. 1995), alors que dans des
3
peuplements matures des taux de mortalité de 70 à 100% ont été observés, tandis ces taux étaient de 30 à 70% pour des peuplements immatures (Maclean 1980). La mortalité des arbres hôtes débute en général quatre à cinq ans après le début de l’épidémie, mais cela varie en fonction du peuplement attaqué et de l’intensité de l’épidémie (Maclean 1980, Morin 1994). La mortalité d’un peuplement est généralement complète 10 ans après le début de l’épidémie (Maclean 1980). Les taux de défoliation et de mortalité sont positivement corrélés à l’abondance de sapin baumier dans le peuplement (Maclean 1980, Royama 1984, Bergeron et al. 1995, Bouchard et al. 2005, Bouchard and Auger 2014), ainsi qu’à l’âge et au diamètre à hauteur de poitrine (DHP) des arbres du peuplement (Maclean 1980, Maclean and Ostaff 1989) ou de la mosaïque forestière (Bergeron et al. 1995). Cependant, la mortalité est négativement corrélée à l’abondance de feuillus du peuplement (Bergeron et al. 1995, Bouchard et al. 2005, Bouchard and Auger 2014). Certains facteurs environnementaux peuvent aussi influencer la défoliation subie par les arbres hôtes, tels que l’élévation (Bouchard and Auger 2014), la température (Baskerville 1975, Bouchard and Auger 2014), et le drainage (Royama 1984).
Une épidémie de la TBE n’entraine pas nécessairement une mortalité complète des arbres. Les individus partiellement défoliés qui survivent à l’épidémie subissent une diminution de leur croissance observable à l’aide de la dendrochronologie (Piene 1980, Maclean 1984, Piene and Little 1990, Bouchard et al. 2005, 2006, Tremblay et al. 2011). Ces mêmes individus parviennent à reconstituer leur feuillage malgré une destruction des bourgeons et une rétro alimentation du vieux feuillage (Piene 1989, Rossi et al. 2009). Pour leur part, les arbres morts lors de l’épidémie laissent place à la régénération préétablie du peuplement qui profite du nouvel apport en lumière et autres ressources induit par les trouées dans la voûte forestière (Tremblay et al. 2011). Dans l’est du Canada, cette régénération préétablie est constituée en grande majorité de sapins, mais aussi d’épinettes, qui sont des espèces tolérantes à l’ombre pouvant vivre sous couvert forestier pendant de nombreuses années, et ce, avec une faible croissance radiale et apicale (Morin 1994, Morin and Laprise 1997, Tremblay et al. 2011). Cette cohorte croît alors de manière marquée et synchrone à la suite d’une épidémie de la TBE (Tremblay et al. 2011). La régénération préétablie d’un peuplement attaqué par cet insecte est d’une grande importance pour l’avenir du peuplement sachant que peu de cônes sont produits par les arbres matures durant une épidémie (Côté and Bélanger 1991). Malheureusement, peu d’études portent spécifiquement sur la régénération préétablie des arbres hôtes attaqués par la TBE, mais surtout, aucune n’a été conduite en temps et lieux lors d’une
4
épidémie. En effet, les études réalisées par Bouchard et al. (2005, 2006), Morin and Laprise (1990), Morin (1994) et Tremblay et al. (2011) se sont basées sur la dendrochronologie afin de reconstituer l’histoire de certains peuplements et sont donc rétrospectives.
Les dommages occasionnés par les épidémies de la TBE recouvrent de très grandes surfaces. Lors de la dernière épidémie, le maximum de défoliation a été observé en 1975 et totalisait une aire de plus de 32 000 000 hectares (MFFP 2016). Quant à elle, l’épidémie ayant lieu depuis 2006 sur la Côte Nord a affecté plus de 6 000 000 ha. Ainsi, l’industrie forestière tente de récupérer rapidement les arbres morts, avant qu’ils ne soient attaqués par des coléoptères xylophages pouvant causer des dommages structuraux au bois tels que le longicorne noir (Monochamus scutellatus; famille des cérambycidés), réduisant ainsi considérablement sa valeur marchande (Raske 1972). Cependant, la coupe de bois entraine elle aussi des conséquences pouvant affecter la régénération. Entre autres, elle peut entraîner une modification de la composition de la forêt boréale au Québec, souvent à la faveur du sapin baumier (Blais 1983, Bouchard and Pothier 2011). Peu d’études au Québec portent sur les répercussions de la récupération du bois après une épidémie de la TBE. Deux études sur la récupération de bois après feu de forêt ont démontré que le passage des machines détruit plus de 70% de la régénération s’étant établi après le passage du feu (Donato et al. 2006, Greene et al. 2006). Les chemins de débardage parcourus par les machines ont d’ailleurs une moins bonne rétention d’eau étant donné qu’ils sont plus compacts (Blouin et al. 2008), et la température du sol y est plus élevée (Fontaine et al. 2010). Notons aussi une étude qui démontre des changements dans la composition (augmentation de l’abondance en feuillus) ainsi que dans l’établissement de semis (diminution comparativement aux peuplement non récoltés) dans des peuplements à dominance de sapins suite à la récolte du bois (Spence and MacLean 2012). Cela est probablement dû au fait que la banque de graines contenue par les individus matures du peuplement est ôtée lors de la récupération, réduisant ainsi les chances d’établissement de semis par la suite (Donato et al. 2006, Greene et al. 2006, Keyser et al. 2009). Une étude portant sur la récupération du bois à la suite d’une épidémie du dendroctone du pin ponderosa (Dendroctonus ponderosae [Erich.]) a démontré que le nombre de semis préétablis était moitié moindre dans les parcelles récoltées comparativement aux parcelles témoins (Griffin et al. 2013). Toutefois, le nombre de semis restants était suffisant pour assurer l’avenir du peuplement. Finalement, l’impact de deux perturbations consécutives, soit l’épidémie de la TBE et la récupération du bois, reste encore incertain quoique très
5
important, et ce, dans un contexte où cela favoriserait l’établissement d’espèces hôtes pour la TBE.
Cette étude vise à comparer l’effet de l’épidémie en termes de défoliation sur la régénération des deux principales espèces hôtes de la région, le sapin baumier et l’épinette noire, tout en identifiant les facteurs influençant cet effet. Aussi, elle vise à comparer la réponse de la régénération du sapin baumier et de l’épinette noire à une épidémie de la TBE avec et sans coupe de récupération. Les objectifs spécifiques sont 1) d’évaluer l’effet de la hauteur et de l’espèce des semis, ainsi que de la composition du peuplement sur leur défoliation, et 2) d’évaluer l’impact de la coupe de récupération sur la défoliation subie par la régénération préétablie. Nous pensons que 1) les individus plus grands devraient être davantage défoliés par la TBE, simplement parce qu’il y a plus de chance qu’une chenille tombe sur un plus gros individu plutôt que sur un plus petit individu. Aussi, les semis de sapin baumier devraient subir davantage de défoliation que les semis d’épinette noire étant donné la plus grande susceptibilité du sapin baumier à la TBE dans la voûte forestière. Pour cette même raison, les peuplements à dominance de sapin baumier devraient avoir attiré davantage de TBE, ce qui signifierait une plus grande défoliation de la régénération préétablie dans ces peuplements. Nous pensons également que 2) la coupe de récupération pourrait influencer la défoliation subie par la régénération préétablie de l’une des deux manières suivantes. D’abord, la coupe pourrait rendre la régénération préétablie plus susceptible aux attaques de la TBE puisque les femelles pourraient pondre davantage dans la strate inférieure (puisque la strate supérieure sera enlevée), et ainsi les chenilles n’auraient que la régénération préétablie pour s’alimenter. Ou alors, la coupe pourrait possiblement épargner la régénération préétablie puisqu’en enlevant les arbres matures constituant la voûte forestière, on enlève les sites de ponte préférés des papillons femelles de la TBE et ainsi l’abondance des chenilles sera moindre dans ces peuplements. Afin de tester ces hypothèses, nous avons évalué la défoliation globale de la régénération préétablie dans 14 sites de peuplements mixtes de sapin baumier et d’épinette noire sur la Côte-Nord (Québec, Canada), dont six sites récoltés entre 2011 et 2013.
6
Chapitre 1
Post-outbreak salvage logging increases defoliation of
black spruce advance regeneration in black
spruce-dominated stands
Anne Cotton-Gagnon
1, Martin Simard
1, Daniel Kneeshaw
2and
Louis De Grandpré
31 Université Laval, Faculté de foresterie, de géographie et de géomatique, Pavillon Abitibi-
Price, Québec, Québec, G1K 7P4, Canada
2 Université de Québec à Montréal, Faculté des sciences, Pavillon des sciences
biologiques, 141 Avenue du Président-Kennedy, Montréal, Québec, H2X 1Y4, Canada
3
Natural Resources Canada, Canadian Forest Service, Laurentian Forestry Centre, 1055 du P.E.P.S., P.O. Box 10380, Stn. Sainte-Foy, Québec, Québec, G1V 4C7, Canada
7
Résumé
Les épidémies de la tordeuse des bourgeons de l’épinette (TBE) sont une perturbation naturelle majeure en forêt boréale nord-américaine et sont souvent suivies de coupes de récupération. Bien que la TBE attaque surtout les arbres matures, elle peut également affecter la régénération pré-établie en sous-couvert. Afin d’évaluer l’impact des facteurs naturels (espèce et hauteur de la régénération, et composition de la canopée) et de la coupe de récupération sur la défoliation subie par la régénération, nous avons échantillonné 14 peuplements défoliés depuis 2006, dont 6 furent récupérés entre 2011 et 2013, dans la région de la Côte-Nord (Québec, Canada). Les résultats démontrent que la régénération de sapin baumier, de grande taille et poussant dans des sapinières subissaient davantage de défoliation. La régénération d’épinette noire dans les pessières était drastiquement plus défoliée dans les sites récupérés que dans les sites naturels, ce qui pourrait influencer la composition future de ces peuplements.
Mots-clés : Tordeuse des bourgeons de l’épinette (Choristoneura fumiferana);
régénération préétablie; sapin baumier (Abies balsamea); épinette noire (Picea mariana); défoliation; coupe de récupération; forêt boréale
8
Abstract
Spruce budworm (SBW) outbreaks are an important natural disturbance in the North American boreal forest, and are often followed by salvage logging. Even though the SBW mainly attacks mature trees, the advance regeneration growing under the canopy may also sustain defoliation. To evaluate the impact of natural factors (regeneration height and species, as well as canopy composition) and of salvage logging on the defoliation sustained by advance regeneration, we sampled 14 stands defoliated since 2006, six of which were salvage logged between 2011 and 2013, on the North Shore region of Québec (Canada). Results show that taller regeneration, balsam fir regeneration, and all regenerating stems in fir-dominated stands sustained more defoliation. Black spruce regeneration growing in black spruce-dominated sites were much more defoliated in salvaged stands than in natural stands, which could lead to changes in future stand composition.
Keywords : Spruce budworm (Choristoneura fumiferana); advance regeneration; balsam fir (Abies balsamea); black spruce (Picea mariana); defoliation; salvage logging, boreal forest
9
1 Introduction
In temperate and boreal biomes, recurrent insect outbreaks are an important component of forest ecosystems, influencing biogeochemical cycles, vegetation dynamics, and resource availability (Hunter 2001, Martin et al. 2006, Edburg et al. 2012). Forest-insect systems have co-evolved for many centuries, leading to a long-term biological resilience of forests to native insect outbreaks, i.e. their ability to absorb change and return to their original state (Drever et al. 2006, Thompson et al. 2009). For example, in almost all outbreak-forming insect taxa (mostly defoliators and bark beetles), the insects primarily attack mature trees, which allows the small understory trees, called advance regeneration, to grow and form the next stand (Mattson and Addy 1975, Morin and Laprise 1997, Greene et al. 1999, Astrup et al. 2008, Boggs et al. 2008, Kayes and Tinker 2012). The spruce budworm (SBW; Choristoneura fumiferana), an insect defoliator native to the North American boreal forest, was even called a super silviculturist (Baskerville 1975) as it kills the overstory layer thus releasing advance regeneration that will form the next stand. However, there are some reports that defoliation of regeneration might occur at high SBW densities, when mature trees are completely defoliated and insects spin down in search of food (Cooke et al. 2007). Given that advance regeneration abundance and quality are critical for post-disturbance replacement of affected stands, an evaluation of the conditions under which advance regeneration is defoliated will improve predictions of post-outbreak stand recovery.
Salvage logging is frequently carried out following forest disturbances such as fire or insect outbreaks (Lindenmayer et al. 2012, Burton et al. 2015). However, post-disturbance regeneration processes differ in many respects between fire and insect outbreaks, which may change the effects of salvage logging on forest recovery. Wildfires severely affect both overstory and understory trees during a relatively short period of time (weeks to months), and in situ regeneration is provided by seeds contained in serotinous cones (conifers; Kozlowski 2012), by suckering and sprouting (hardwoods; Bergeron and Charron 1994) or by aerial seed dispersion from undisturbed sites (Greene et al. 1999). In contrast, insect outbreaks primarily affect overstory trees, are species-specific and tend to last several years, during which recruitment by seeds is low (Batzer and Popp 1985, Astrup et al. 2008, Man and Rice 2010, Moulinier et al. 2011), emphasizing the importance of pre-established advance regeneration. These inherent differences between disturbances characteristics may determine salvage logging effects on forest regeneration processes, and consequently, future stand composition. For example, a compositional shift
10
could occur following SBW outbreaks (Burton et al. 2015) if salvage logging increases the vulnerability of the advance regeneration by concentrating defoliation in the regeneration layer. The direction or quantity of change in forest succession following salvage logging remains largely unresolved because of the diversity of forest-insect systems, the complexity of their regeneration processes, and the scarcity of studies on salvage logging, especially after insect outbreaks.
Defoliation of advance regeneration could be affected by at least three factors. First, larger understory trees should be more defoliated than smaller ones because larvae have a greater probability of falling onto large stems than onto small ones. Second, the different species (balsam fir [Abies balsamea (L.) Mill] vs. black spruce [Picea mariana (Mill.) B.S.P]) of advance regeneration could experience differing defoliation levels because larvae spinning down from the canopy in the spring may arrive at a moment when budburst has occurred in balsam fir but not in black spruce or, alternatively, if down-spinning occurs later in the season, then both hosts may be equally vulnerable. Earlier budburst in balsam fir explains a large part of the increased vulnerability of fir compared to black spruce in mature trees. However, due to the timing of larvae arrival, differential defoliation among host species in the understory might be different from that observed in the canopy. Third, stands with a higher percentage of balsam fir in the canopy should have higher SBW populations than in black spruce-dominated stands, increasing the probability of larvae falling on advance regeneration in fir-dominated stands.
Salvage logging could either protect advance regeneration or increase its risk of being defoliated by the SBW. Salvage logging could reduce the defoliation of advance regeneration because removal of the overstory would reduce the abundance of larvae spinning down and feeding on saplings and seedlings. Alternatively, salvage logging could increase defoliation of advance regeneration because the remaining saplings and seedlings would be the only egg-laying sites for SBW moths and subsequently the only food available for the larvae.
The general objective of this study was to acquire insight into the regeneration processes of a boreal forest affected by a SBW outbreak, and how salvage logging influences those processes. Specifically, we aimed (1) to determine the effects of height and species of advance regeneration and of canopy composition on the severity of the defoliation sustained by advance regeneration and (2) to determine the effects of salvage logging on the defoliation of advance regeneration. For the first objective, we hypothesized that (1) taller individuals would be more defoliated than smaller ones, that balsam fir regeneration
11
would sustain more defoliation than black spruce regeneration, and that advance regeneration in balsam fir-dominated stands would experience more defoliation than in mixed or spruce-dominated stands. For the second objective, we considered two alternative hypotheses, i.e., that advance regeneration located in salvage logged stands would either sustain less or more defoliation than that located in non-harvested stands.
12
2 Methods
2.1 Study area
A SBW outbreak which started in 2006 in the eastern boreal forest of Quebec (Canada), along with ongoing salvage logging operations in that area, provided the perfect opportunity to address these questions. The study area is located on the North Shore of the St. Lawrence River and east of the Saguenay fjord in Quebec. The arctic and sub-humid climate is characterized by a mean annual temperature of 1.7°C (mean January temperature: -14.3°C and mean July temperature: 15.6°C), and mean annual precipitation of 1040.5 mm, 33% of which falls as snow (Environnement Canada 2016). The landscape is characterised by tall hills that rarely exceed 500 m, some rock cliffs and many valleys, while the metamorphic bedrock is covered mainly by thin tills. Forests are characterized by a mixture of balsam fir and black spruce, the latter increasing in abundance northward and inland (Robitaille and Saucier 1998, De Grandpré et al. 2009).
2.2 Site selection
Eight sites, hereafter referred to as “natural sites”, were selected in old uneven-aged stands (120+ years) defoliated by the SBW but not harvested, representing a composition gradient ranging from balsam fir-dominated to black spruce-dominated stands. Six sites, hereafter referred to as “harvested sites”, were selected in stands that were salvage logged in 2011 (one stand), in 2012 (four stands), or in 2013 (one stand). In all sites, salvage harvest was carried out using the careful logging norms (harvesting with protection of regeneration and soils or HPRS), which is the main type of logging in Quebec’s forests. Prior to harvesting, these stands were similar to the natural sites (i.e., old uneven-aged stands growing on till deposits) and their composition was selected to have a codominance of balsam fir and black spruce. All sites were at least 25 m from a road and showed no sign of other disturbances. To verify that there were no differences between the defoliation history in the natural and harvested sites, we used annual aerial detection survey data to compute cumulative defoliation in each stand (MFFP 2015). Starting in 2006 (the first year of the outbreak), we added the defoliation values (1 = low, 2 = moderate, and 3 = severe) to obtain a defoliation severity index (Simard and Lajeunesse 2015) representing cumulative defoliation since the beginning of the outbreak.
13
2.3 Sampling
2.3.1 Advance regeneration
During the summer of 2015, one (in natural sites) or two (in harvested sites) 100-m long transects were sampled at each site to evaluate the abundance, composition and defoliation of advance regeneration. Natural sites were installed in 2013 whereas harvested sites were installed in 2015. Two transects were installed in the harvested sites to account for spatial heterogeneity due to skid trails and unsalvaged patches. At each site, 60 live balsam fir and 60 live black spruce regeneration stems were randomly selected along the transects, in each of the five following height classes (total of 300 stems per species): 8 to 14.9 cm, 15 to 49.5 cm, 50 cm to 1.29 m, 1.3 to 2.29 m, and > 2.3 m but < 5 cm dbh (diameter at breast height; 1.3 m above the ground). We tagged and mapped each stem, and noted their status (live or dead; all live stems in 2013 in natural sites and in 2015 in harvested sites) and cumulative defoliation caused by the SBW according to the following categories: 0 to 5%, 6 to 25%, 26 to 50%, 51 to 75%, 76 to 95%, and 96 to 100%.
At each site, more precise measurements were made of a subsample of 10 stems of each species and of each height class, for a total of 50 balsam fir stems and 50 black spruce stems. The exact height (for the first four size classes) or dbh (for the fifth size class) of these stems were measured, as well as the width of their crown where the branches were the longest. For stems in the fifth height class (>2.3 m tall but < 5 cm dbh), dbh was converted to height using allometric equations developed on a sample of 144 balsam fir and 74 black spruce saplings (balsam fir: R2 = 0.70; black spruce: R2 = 0.75).
14 2.3.2 Mature trees
In natural sites, all living and dead trees were identified and mapped in a 40 x 100-m quadrat, and their crown width and dbh were measured. In harvested sites, pre-logging stand composition was reconstructed by tallying all live trees, standing dead trees and cut stumps in two 2 m x 100-m transects positioned on the regeneration transects, for a total of 400 m2 per site. Species, status, decay class and dbh were noted for all trees, whereas species (determined based on bark left on the stumps), decay class, cause of death (harvested or not), and basal diameter were noted for all stumps. Basal diameters were converted to diameter at breast height using allometric relationships developed from 104 balsam fir trees and 98 black spruce trees (balsam fir: R2 = 0.97; black spruce: R2 = 0.98). Pre-logging basal area per species was computed using live trees, recently dead trees (decay class of 4 or less), and harvested stumps, all with a dbh ≥ 9 cm.
2.4 Statistical analysis
To assess the effects of species and height of advance regeneration, stand-scale and neighbourhood composition (see below), harvest, and number of years since harvest on cumulative defoliation of advance regeneration, we used mixed-effects linear models in R (version 3.2.2; R Core Team 2015). This function allowed the models to account for environmental differences among sites, which were treated as a random factor. To meet our objectives, two separate regressions were performed: one using natural sites only (n = 2195 regeneration stems), and the other combining all natural and harvested sites (n = 7577 stems). In all cases, the median of each regenerating stem’s defoliation class was used as the response variable and site was considered as a random variable. Because height classes were available for the whole sample, were used this variable instead of exact heights or crown widths in the models.
Because SBW larvae fall from the canopy onto understory stems, the defoliation of a given sapling or seedling may be influenced by the composition of the canopy directly above the stem. Therefore, the effect of canopy composition was assessed both at the stand scale (whole site) and at the local scale, i.e. in the stems’ neighbourhoods (in a 5-meters radius around each regenerating stem) for natural sites. For harvested sites, only pre-logging stand-scale composition was used to assess its effect on regeneration defoliation. Stand-level canopy composition was calculated as the percentage of balsam fir basal area relative to total basal area of all tree species in the site. Neighbourhood composition was
15
determined for each stem of advance regeneration by overlaying the maps of georeferenced mature trees and sampled advance regeneration in ArcGIS (ESRI 2012). First, we created a 5-m radius buffer around each regeneration stem to represent the local composition in their neighbourhoods. Second, we created buffers representing the horizontal extent of the crown of each mature tree using species-specific allometric relationships between dbh and crown width (balsam fir: R2 = 0.50, P < 0.00001, n = 1044; black spruce: R2 = 0.28, P < 0.00001, n = 2169). The two buffer layers were then intersected to calculate, for each regenerating stem, the proportion of balsam fir, black spruce, white spruce (Picea glauca (Moench) Voss), and open space in the canopy in a 5-m radius. Neighbourhood co5-mposition was calculated as the percentage of the area occupied by balsam fir over the total area of the advance regeneration’s surroundings, including the area unoccupied by canopy trees.
16
3 Results
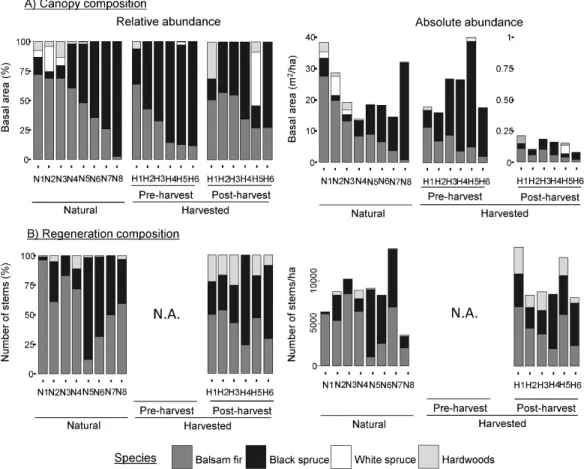
3.1 Stand composition and defoliation history
Relative balsam fir basal area in the canopy ranged from 2.6% to 72.2% in natural sites, from 11.8% to 63.9% in harvested sites prior to salvage logging and from 27.3% to 57.0% following salvage logging (Fig. 1). Other tree species in the canopy were mostly black spruce, along with some white spruce and occasional hardwood species. Defoliation history was variable but comparable between the natural and harvested sites, except for one of the natural sites (site N3) that had 5 to 10% more defoliation than the others (Fig. 2). Thus, any differences between the defoliation of advance regeneration in natural sites versus harvested sites should not be a result of differences in defoliation history.
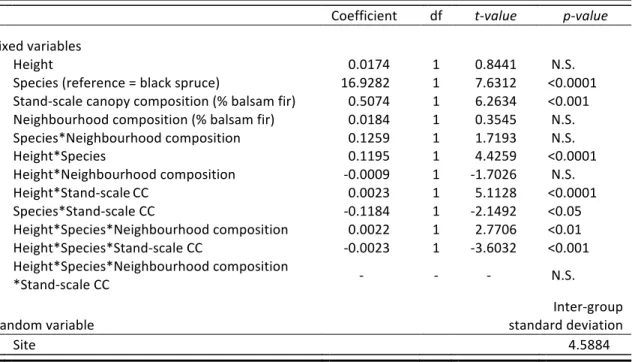
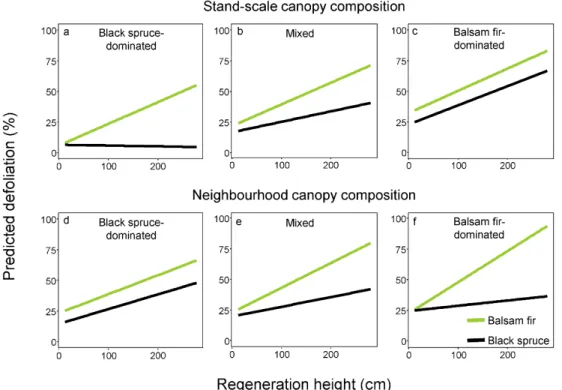
3.2 Factors influencing defoliation of advance regeneration in natural sites
Defoliation increased with regeneration stem height and this relationship differed significantly by species and with stand-scale canopy composition (three-way interaction; however the four-way interaction, i.e., with the neighbourhood composition, was not significant (Fig. 3, Tab. 1)). Balsam fir was consistently more defoliated than black spruce (17% more defoliation; Tab. 1), and the positive relationship between regeneration height and defoliation increased as the proportion of balsam fir in the canopy increased (Fig. 3a, b and c). Specifically, defoliation of black spruce regeneration in black spruce-dominated stands was almost zero and did not change with stem height (Fig. 3a), but as the proportion of balsam fir increased in the canopy, black spruce defoliation also increased, especially for larger stems (Fig. 3a, b and c).
Neighbourhood (5-m radius) canopy composition had a significant but weaker effect (Height * Species * Neighbourhood CC: P < 0.01; Tab. 1) than stand-scale canopy composition (Height * Species * Stand-scale CC: P < 0.001; Tab. 1). Whereas the relationships between defoliation and canopy composition were similar at the neighbourhood vs. stand scale in mixed composition stands (Fig. 3b and e), they were different in pure compositions for black spruce regeneration. Notably, at the local scale, black spruce regeneration was more defoliated in black spruce-dominated neighbourhoods (16 % to 47%; Fig. 3d) than in balsam fir-dominated neighbourhoods (25% to 36%; Fig. 3f), whereas the opposite was observed at the stand scale.
17
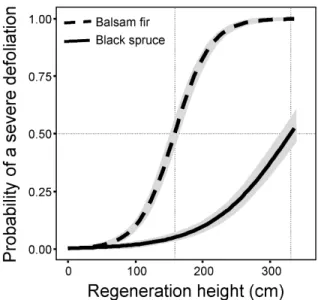
To determine the height above which regenerating stems were most likely to be severely defoliated, we used a logistic regression between defoliation severity (light : defoliation < 50% vs. severe : defoliation ≥ 50%) and stem height (Fig. 4). Balsam fir stems > 120 cm had a greater (Cote et al. 2014) probability of sustaining severe defoliation and the probability reached 0.95 above 2 m in height. For black spruce, the probability of sustaining severe defoliation increased (Cote et al. 2014) only in saplings taller than 3.5 m. Using the exact height or crown width (measured in the subsample), instead of height class medians, gave identical results. Therefore, using height classes, which saves times in the field and is less prone to variability among measurers, is sufficient for assessing advance regeneration vulnerability (height classes vs. exact height: r = 0.97; height classes vs. crown width: r = 0.85; exact height vs. crown width: r = 0.88; n = 675; data not shown otherwise).
3.3 Effect of salvage harvest on defoliation of advance regeneration
Time since harvest did not affect defoliation of advance regeneration (P = 0.2919). The model combining natural and harvested sites showed that in both harvested sites and natural sites, defoliation was greater for balsam fir regeneration and increased with regeneration height and with the proportion of balsam fir in the stand-scale canopy composition (Fig. 5, Tab. 2). However, in stands that were dominated by black spruce before logging, the effect of harvesting greatly increased defoliation of black spruce regeneration after harvesting (P < 0.0001; Tab. 2, Fig. 5a). Whereas defoliation of tall (> 2.5 m) black spruce regeneration reached 30% in harvested sites, this defoliation did not even reach 10% in natural sites (Fig. 5a).
18
4 Discussion
4.1 Factors influencing defoliation of regeneration in natural sites
SBW outbreaks have affected the boreal forest for centuries, shaping its dynamics and regeneration processes (Morin 1994), yet little is known about the effects of defoliation of advance regeneration. This study provides insight into how SBW outbreaks affect different size classes of regeneration of balsam fir and black spruce in natural and salvage logged sites.
First, defoliation increased with height of advance regeneration (i.e., slopes were positive; Fig. 3), which supports our first hypothesis. This can be explained by the fact that taller stems have wider crowns, and thus a greater chance of intercepting larvae spinning down from the canopy. Smaller regenerating stems may also benefit from the sheltering effect provided by taller stems, which would intercept falling larvae and in turn reduce the probability of larvae landing on smaller stems. Ruel and Huot (1993) found similar results, as tall balsam fir regeneration (> 2 m) was practically absent from balsam fir-dominated stands, where SBW defoliation lasted the longest. Similar to the gyspy moth (Lymantria dispar dispar Linnaeus) -forest system (Hix et al. 1991), succession in attacked stands may be ensured by the smaller seedlings already in place, as tall advance regeneration sustains greater defoliation and mortality.
Also consistent with our hypothesis, balsam fir regeneration was more defoliated than black spruce regeneration (Fig. 3 and Fig. 4). As in the overstory, balsam fir regeneration is more vulnerable than black spruce, which may be due to better phenological synchrony between larval emergence and bud opening in balsam fir (Maclean 1980, Nealis and Régnière 2004). However, if down-spinning occurs after black spruce buds are open, the greater vulnerability of balsam fir may also be due to (i) black spruce’s greater foliage biomass, from approximately 25% to 50% more than balsam fir for the studied range of DBH, which allows black spruce to better sustain the defoliation caused by a given density of larvae (Lambert et al. 2005, Ung et al. 2008), (ii) the greater propensity of black spruce to produce epicormic shoots, allowing the replacement of consumed foliage (Simard and Payette 2003), or (iii) differences in foliage quality (nutrient contents, chemical and/or physical defenses, etc.) between the two studied species factors (Blais 1957, Hudes and Shoemaker 1984, Simard and Payette 2003, Monerris et al. Accepted, Lambert et al. 2005, Ung et al. 2008), or some combination of all these.
19
Seedling defoliation also increased as the proportion of balsam fir in the canopy increased, which supports our third hypothesis (Fig. 3). For black spruce seedlings defoliation, this effect was stronger in fir-dominated stands than in fir-dominated neighbourhoods (Fig. 3a). The stronger relationship in fir-dominated stands vs neighbourhoods may be due to SBW population density such that high densities in balsam fir-dominated stands may lead to greater defoliation of all stems in the understory. Whereas in neighbourhoods, ballooning of down-spinning threads may lead to larvae not descending directly below infested trees. Bognounou et al. (2017) also suggest that there may be temporal variability in defoliation to neighbourhoods and stand-level defoliation patterns between species.
4.2 Effect of harvest on regeneration defoliation
Salvage logging did not qualitatively change the relationships found in natural sites, except for one notable case: in black spruce-dominated stands, salvage logging led to greater defoliation of tall black spruce regeneration, thus removing beneficial protective effects of the black spruce overstory (Fig. 5a). The immediate effect of logging was to cause the understory to become the canopy, which will also lead to a shift from a black spruce-dominance to a mixed composition, as there is usually a high proportion of balsam fir in the understory (Fig 1). This shift in composition is consistent with the post-harvest shift observed by Burton et al. (2015) and may lead to the associational susceptibility of black spruce regeneration (Atsatt and O’Dowd 1976).
The present study however demonstrates that the natural regeneration processes of a boreal forest attacked by the SBW may be compromised by salvage logging in black spruce-dominated stands. Under natural dynamics, black spruce is maintained during outbreaks because of the preference of SBW for balsam fir, and as a corollary, stands in which black spruce is abundant diminish the future impact of the SBW. Hix et al. (1991) found similar results as oak (Quercus spp.) forests attacked by the gypsy moth underwent an increase in ligneous species diversity, which may reduce their susceptibility to future attacks by that insect. Following harvesting of black spruce stands, however, regeneration processes are compromised as the overstory that is removed is much less vulnerable than fir (black spruce has 28% as much defoliation as balsam fir; Hennigar et al. 2008) while the opening of the canopy exposes black spruce regeneration to defoliation (30% defoliation in harvested sites versus 5% in natural sites) six times higher than in the understory. Shifting composition away from black spruce consequently increases forests susceptibility to future infestations by the SBW.
20
4.3 Implications for sustainable forest management
Similar to results found on salvage logging following a spruce beetle (Dendroctonus rufipennis Kirby) outbreak, this study demonstrates how forest management should be locally adapted in response to large-scale infestations (Boucher and Mead 2006). The increased defoliation of black spruce regeneration in salvage-logged black spruce-dominated stands might reduce black spruce recruitment and lead to an increase in balsam fir in the long term, especially since balsam fir is a highly competitive shade-tolerant species (Messier et al. 1999). This increase in the proportion of balsam fir coupled to a decrease in species diversity could then lead to a forest more susceptible to future SBW outbreaks. Long-term studies are needed to confirm this trend, but changes in salvage logging practices should be implemented immediately to secure the black spruce component of the boreal forests and reduce their overall susceptibility to the SBW.
21
Acknowledgements
We thank Gauthier Cervello, Hugues Dorion, Mathieu Gauvin and Jeanne Moisan-Perrier for field support as well as Dave Gervais and Stéphane Bourassa (CFS-NRCan) for technical support and experiment design. We also express gratitude to Maryse Marchand for statistical assistance. At last, we thank the Laurentian Forestry Centre for the equipment and workplace. Financial support was provided by the Fonds de recherche Nature et Technologies of Québec.
22
References cited
Astrup, R., K. D. Coates, and E. Hall. 2008. Recruitment limitation in forests: lessons from an unprecedented mountain pine beetle epidemic. Forest Ecology and Management 256:1743-1750.
Atsatt, P. R. and D. J. O’Dowd. 1976. Plant defense guilds. Science 193:24-29.
Baskerville, G. L. 1975. Spruce budworm - super silvi-culturist. Forestry Chronicle 51:138-140.
Batzer, H. O. and M. P. Popp. 1985. Forest succession following a spruce budworm outbreak in Minnesota. Forestry Chronicle 61:75-80.
Bergeron, Y. and D. Charron. 1994. Postfire stand dynamics in a southern boreal forest (Quebec): A dendroecological approach. Ecoscience 1:173-184.
Blais, J. 1957. Some relationships of the spruce budworm, Choristoneura fumiferana (Clem.) to black spruce, Picea mariana (Moench) Voss. Forestry Chronicle 33:364-372.
Boggs, K., M. Sturdy, D. J. Rinella, and M. J. Rinella. 2008. White spruce regeneration following a major spruce beetle outbreak in forests on the Kenai Peninsula, Alaska. Forest Ecology and Management 255:3571-3579.
Bognounou, F., L. De Grandpré, D. S. Pureswaran, and D. Kneeshaw. 2017. Temporal variation in plant neighborhood effects on the defoliation of primary and secondary hosts by an insect pest. Ecosphere 8.
Boucher, T. V. and B. R. Mead. 2006. Vegetation change and forest regeneration on the Kenai Peninsula, Alaska following a spruce beetle outbreak, 1987–2000. Forest Ecology and Management 227:233-246.
Burton, P. J., M. Svoboda, D. Kneeshaw, and K. W. Gottschalk. 2015. Options for promoting the recovery and rehabilitation of forests affected by severe insect outbreaks. Restoration of Boreal and Temperate Forests:495.
23
Cooke, B. J., V. G. Nealis, and J. Régnière. 2007. Insect defoliators as periodic
disturbances in northern forest ecosystems. Plant disturbance ecology: the process and the response. Edited by EA Johnson and K. Miyanishi. Elsevier Academic Press, Burlington, Mass., USA:487-525.
Cote, D., C. Lupi, R. Gagnon, D. Lord, and H. Morin. 2014. Growth dynamics of
successive post-fire cohorts of black spruce: Is site potential reduced? (vol 90, pg 96, 2014). Forestry Chronicle 90:135-135.
De Grandpré, L., S. Gauthier, C. Allain, D. Cyr, S. Périgon, A. Pham, D. Boucher, J. Morissette, G. Reyes, and T. Aakala. 2009. Towards an ecosystem approach to managing the boreal forest in the North Shore region: disturbance regime and natural forest dynamics. Ecosystem management in the boreal forest. Edited by S. Gauthier, M.-A. Vaillancourt, A. Leduc, L. De Grandpré, D. Kneeshaw, H. Morin, P. Drapeau, and Y. Bergeron. Presses de l'Université du Québec, Montreal, Quebec, Canada:229-255.
Drever, C. R., G. Peterson, C. Messier, Y. Bergeron, and M. Flannigan. 2006. Can forest management based on natural disturbances maintain ecological resilience? Canadian Journal of Forest Research 36:2285-2299.
Edburg, S. L., J. A. Hicke, P. D. Brooks, E. G. Pendall, B. E. Ewers, U. Norton, D. Gochis, E. D. Gutmann, and A. J. Meddens. 2012. Cascading impacts of bark beetle-caused tree mortality on coupled biogeophysical and biogeochemical processes. Frontiers in Ecology and the Environment 10:416-424.
Environnement Canada. 2016. Canadian climate normals 1981-2010. URL : www.climate.weather.gc.ca/climate_normals/. Accessed November 10, 2016. ESRI. 2012. ArcGIS Desktop: Release 10. Redlands, CA: Environmental Systems
Research Institute.
Greene, D. F., J. C. Zasada, L. Sirois, D. Kneeshaw, H. Morin, I. Charron, and M. J.
Simard. 1999. A review of the regeneration dynamics of North American boreal forest tree species. Canadian Journal of Forest Research-Revue Canadienne De
24
Hennigar, C. R., D. A. MacLean, D. T. Quiring, and J. A. Kershaw. 2008. Differences in spruce budworm defoliation among balsam fir and white, red, and black spruce. Forest Science 54:158-166.
Hix, D. M., D. E. Fosbroke, R. R. Hicks Jr, and K. W. Gottschdk. 1991. Development of regeneration following gypsy moth defoliation of Appalachian Plateau and Ridge & Valley hardwood stands. 8th Central Hardwood Forest Conference.
Hudes, E. S. and C. A. Shoemaker. 1984. Statistical comparison of spruce budworm (Lepidoptera: Tortricidae) development on balsam fir and on red and black spruce. Environmental Entomology 13:1602-1603.
Hunter, M. D. 2001. Insect population dynamics meets ecosystem ecology: effects of herbivory on soil nutrient dynamics. Agricultural and Forest Entomology 3:77-84. Kayes, L. J. and D. B. Tinker. 2012. Forest structure and regeneration following a
mountain pine beetle epidemic in southeastern Wyoming. Forest Ecology and Management 263:57-66.
Kozlowski, T. T. 2012. Fire and ecosystems. Elsevier.
Lambert, M., C. Ung, and F. Raulier. 2005. Canadian national tree aboveground biomass equations. Canadian Journal of Forest Research 35:1996-2018.
Lindenmayer, D. B., P. J. Burton, and J. F. Franklin. 2012. Salvage logging and its ecological consequences. Island Press.
Maclean, D. A. 1980. Vulnerability of fir-spruce stands during uncontrolled spruce budworm outbreaks - a review and discussion. Forestry Chronicle 56:213-221.
Man, R. and J. A. Rice. 2010. Response of aspen stands to forest tent caterpillar defoliation and subsequent overstory mortality in northeastern Ontario, Canada. Forest Ecology and Management 260:1853-1860.
Martin, K., A. Norris, and M. Drever. 2006. Effects of bark beetle outbreaks on avian biodiversity in the British Columbia interior: Implications for critical habitat management. Journal of Ecosystems and Management 7.
25
Mattson, W. J. and N. D. Addy. 1975. Phytophagous insects as regulators of forest primary production. Science 190:515-522.
Messier, C., R. Doucet, J. C. Ruel, Y. Claveau, C. Kelly, and M. J. Lechowicz. 1999. Functional ecology of advance regeneration in relation to light in boreal forests. Canadian Journal of Forest Research 29:812-823.
MFFP. 2015. Données sur les perturbations naturelles - insectes : Tordeuse des bourgeons de l'épinette. Québec, gouvernement du Québec, Direction de la protection des forêts. URL: www.donneesquebec.ca/recherche/fr/dataset/donnees-sur-les-perturbations-naturelles-insecte-tordeuse-des-bourgeons-de-lepinette. Accessed June 13, 2016.
Monerris, J., L. De Grandpré, D. S. Pureswaran, H. Morin, and D. Kneeshaw. Physical and nutritional properties of foliage of two hosts of eastern spruce budworm. Accepted in Forests.
Morin, H. 1994. Dynamics of balsam fir forests in relation to spruce budworm outbreaks in the boreal zone of Quebec. Canadian Journal of Forest Research 24:730-741.
Morin, H. and D. Laprise. 1997. Seedling bank dynamics in boreal balsam fir forests. Canadian Journal of Forest Research 27:442-1451.
Moulinier, J., F. Lorenzetti, and Y. Bergeron. 2011. Gap dynamics in aspen stands of the Clay Belt of northwestern Quebec following a forest tent caterpillar outbreak. Canadian Journal of Forest Research 41:1606-1617.
Nealis, V. G. and J. Regniere. 2004. Insect-host relationships influencing disturbance by the spruce budworm in a boreal mixedwood forest. Canadian Journal of Forest Research 34:1870-1882.
R Core Team. 2015. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/.
26
Robitaille, A. and J. Saucier. 1998. Paysages régionaux du Québec méridional.
Gouvernement du Québec, Ministére des ressources naturelles. Les Publications du Quebec, Quebec.
Ruel, J.-C. and M. Huot. 1993. Influence of the spruce budworm, choristoneura-fumiferana (clem), on regeneration of fir stands after clearcutting. Forestry Chronicle 69:163-172.
Simard, M. and P. Lajeunesse. 2015. The interaction between insect outbreaks and debris slides in a glacial valley of the Eastern Canadian Shield. Ecosystems 18:1281-1289.
Simard, M. and S. Payette. 2003. Accurate dating of spruce budworm infestation using tree growth anomalies. Ecoscience 10:204-216.
Thompson, I., B. Mackey, S. McNulty, and A. Mosseler. 2009. Forest resilience, biodiversity, and climate change. Page 67 in A synthesis of the
biodiversity/resilience/stability relationship in forest ecosystems. Secretariat of the Convention on Biological Diversity, Montreal. Technical Series.
Ung, C.-H., P. Bernier, and X.-J. Guo. 2008. Canadian national biomass equations: new parameter estimates that include British Columbia data. Canadian Journal of Forest Research 38:1123-1132.
27
Tables
Table 1 Results of the mixed model explaining cumulative defoliation (%) of advance regeneration in natural sites (n = 2195). N.S. = not significant (alpha = 5%); df = degrees of freedom; CC = canopy composition.
Coefficient df t-value p-value
Fixed variables Height 0.0174 1 0.8441 N.S. Species (reference = black spruce) 16.9282 1 7.6312 <0.0001 Stand-scale canopy composition (% balsam fir) 0.5074 1 6.2634 <0.001 Neighbourhood composition (% balsam fir) 0.0184 1 0.3545 N.S. Species*Neighbourhood composition 0.1259 1 1.7193 N.S. Height*Species 0.1195 1 4.4259 <0.0001 Height*Neighbourhood composition -0.0009 1 -1.7026 N.S. Height*Stand-scale CC 0.0023 1 5.1128 <0.0001 Species*Stand-scale CC -0.1184 1 -2.1492 <0.05 Height*Species*Neighbourhood composition 0.0022 1 2.7706 <0.01 Height*Species*Stand-scale CC -0.0023 1 -3.6032 <0.001 Height*Species*Neighbourhood composition *Stand-scale CC - - - N.S. Random variable Inter-group standard deviation Site 4.5884
28
Table 2 Results of the mixed model explaining cumulative defoliation (%) of advance regeneration in natural and harvested sites (n = 7577). N.S. = not significant (alpha = 5%); df = degrees of freedom. In this analysis, canopy composition was only computed at the stand scale.
Coefficient df t-value p-value
Fixed variables Time since harvest - - - N.S. Height -0.0091 1 -0.6448 N.S. Species (reference = black spruce) 21.8487 1 13.7301 <0.0001 Canopy composition (stand-scale; % balsam fir) 0.5795 1 9.4194 <0.0001 Logging 14.6819 1 3.3302 N.S. Height*Species 0.1432 1 7.9457 <0.0001 Height*Canopy composition 0.0018 1 6.7931 <0.0001 Species*Canopy composition 0.1231 1 7.3090 <0.0001 Height*Species*Canopy composition -0.1301 1 -4.2580 <0.001 Logging*Height -18.7210 1 -9.2119 <0.0001 Logging*Species -0.3963 1 -3.8151 <0.0001 Logging*Canopy composition -0.0013 1 -3.6283 N.S. Logging*Height*Species -0.1303 1 -5.8460 <0.0001 Logging*Species*Canopy composition 0.3318 1 7.0443 <0.0001 Logging*Height* Species*Canopy composition 0.0026 1 5.1479 <0.0001 Random variable standard deviationInter-group
29
Figures
Figure 1 Relative (left) and absolute (right) stand composition of the canopy (top) and regeneration (bottom) for the natural (N) and harvested (H) sites. N.A. = not available.
30
Figure 2 Cumulative defoliation history in natural sites (circles) versus harvested sites (triangles). Open symbols indicate superposed trajectories.
31
Figure 3 Predicted defoliation of advance regeneration according to height and species of regeneration for three stand-scale canopy compositions (top; in black spruce-dominated stands, the relative basal area of balsam fir was less than 2%, averaged 42% in mixed stands, and was greater than 72% in balsam fir-dominated stands), and three neighbourhood compositions (bottom; black spruce-dominated neighbourhoods have a relative basal area of balsam fir that is 10%, that averages 50% for mixed neighbourhoods, and is greater than 90% for balsam fir-dominated neighbourhoods). Stand-scale composition graphs have been calculated based on the mean value of neighbourhood compositions and vice versa.
32
Figure 4 Probability of sustaining severe (≥ 50%) defoliation as a function of stem height. Shaded areas indicate the 9th confidence interval.
33
Figure 5 Predicted defoliation of advance regeneration in natural and harvested sites by stem height for the two studied species and three stand-scale canopy compositions (for black spruce-dominated stands relative basal area in balsam fir is 10%, it is 40% for mixed stands, and 65% for balsam fir-dominated stands).