Caractérisation structurale et de liaison
membranaire de rétinol déshydrogénases
Thèse
Mustapha Lhor
Doctorat en biologie cellulaire et moléculaire
Philosophiae doctor (Ph. D.)
Québec, Canada
Caractérisation structurale et de liaison
membranaire de rétinol déshydrogénases
Thèse
Mustapha Lhor
Sous la direction de :
Résumé
Les rétinol déshydrogénases ou RDHs sont des oxydoréductases inhérentes à l’accomplissement de la fonction visuelle de la rétine. Elles sont en effet impliquées dans le cycle visuel rétinien. Suite à l’absorption de la lumière par le pigment visuel des photorécepteurs, la rhodopsine, la RDH8 est la première enzyme qui va intervenir dans le cycle visuel après la libération du chromophore de la rhodopsine, le tout-trans rétinal. Ainsi, la RDH8 détoxifie les photorécepteurs car le tout-trans rétinal est une espèce très réactive qui peut induire des dommages à la rétine. La RDH11, quant à elle, agit de concert avec la RDH5 au niveau de la dernière étape du cycle visuel dans l’épithélium pigmentaire rétinien en transformant le 11-cis rétinol en 11-cis rétinal, qui sera réacheminé vers les photorécepteurs pour régénérer le pigment visuel. Toutefois, la structure tertiaire des RDHs n’a encore jamais été résolue. Ces enzymes sont néanmoins reconnues pour être associées aux membranes cellulaires par leur segment N- et/ou C-terminal. Nous avons alors entrepris ce travail afin de caractériser la structure de ces enzymes et mieux comprendre leur interaction avec les membranes. Nous avons étudié dans un premier temps différentes portions du segment N- et C-terminal de la RDH11 et la RDH8 respectivement, par différentes méthodes spectroscopiques. Nous avons alors observé que les segments de ces deux enzymes agissent par deux modes d’action totalement différents. La RDH11 ferait appel à un segment N-terminal transmembranaire qui adopte une conformation hélicale peu importe sa longueur, alors que la RDH8 utiliserait un segment C-terminal qui adopte une structure secondaire variable selon la longueur et dont la liaison est périphérique à la membrane. En plus, la liaison de la RDH8 par son segment C-terminal serait potentiellement facilitée par une ou plusieurs acylations situées au niveau de certaines cystéines. Les mesures de pression d’insertion maximale ont permis de comparer les interactions entre des segments de longueur variable en N-terminal de la RDH11 et en C-terminal de la RDH8 avec des monocouches de différents phospholipides. Ainsi, nous avons déterminé les interactions les plus favorables pour chacun de ces segments.
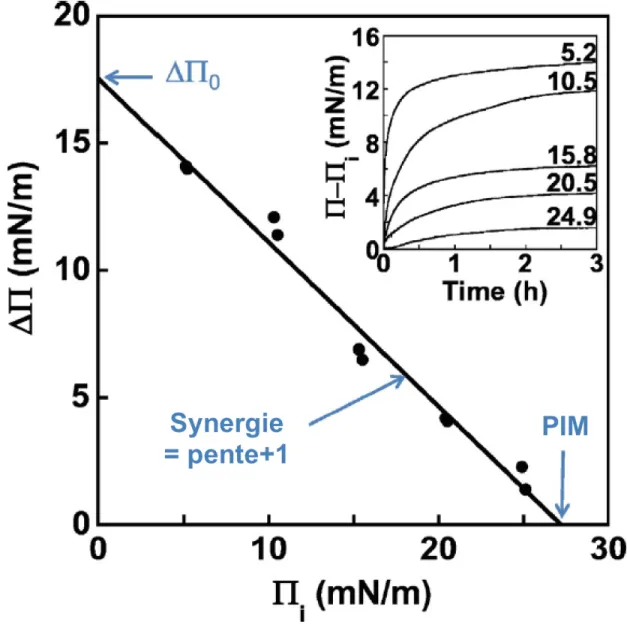
Nous nous sommes focalisés par la suite sur l’étude de l’enzyme RDH8 et la comparaison de ses propriétés structurales, de stabilité et de liaison membranaire avec celles de sa forme tronquée RDH8t, dépourvue de son segment en C-terminal. Notons que nous avons mis au point un protocole adapté pour surexprimer et purifier la RDH8 et sa forme tronquée. À notre connaissance, il s’agit ici des premiers travaux de recherche rapportant la surexpression et la purification d’une RDH8 (bovine) complète dans un système procaryote (E. coli). Nous avons alors constaté que les deux formes de la RDH8, complète et tronquée, comprenaient majoritairement des hélices α en plus de la présence de feuillets β, en accord avec le motif de Rossmann fold suggéré dans la littérature pour cette famille d’enzymes. Il s’est avéré également que le segment C-terminal a un impact sur la stabilité de la RDH8 comme démontré par les mesures du contenu en structure secondaire de ces protéines en fonction des conditions de stockage et dans les expérimentations de dénaturation thermique. Enfin, les mesures de pression d’insertion maximale (PIM) et de synergie ont démontré que le segment C-terminal facilitait la liaison membranaire de la forme complète par rapport à la forme tronquée, notamment dans le contexte de phospholipides portant une tête polaire chargée négativement. L’interaction membranaire de la RDH8 pourrait donc impliquer des interactions électrostatiques. Des expériences de spectroscopie de fluorescence ont permis de confirmer l’implication du segment C-terminal dans la liaison de la RDH8 avec des bicouches lipidiques grâce à la présence de deux résidus tryptophanes uniquement dans son segment C-terminal.
Abstract
In the retina, retinol dehydrogenases (RDHs) play a crucial role in the visual cycle allowing a good vision. The first step of the visual cycle is taking place in photoreceptors where RDH8 is located and then in the retinal pigmented epithelium (RPE) where RDH11 can be found.
RDH11 is likely anchored to membranes by means of its N-terminal segment whereas RDH8 has been postulated to be membrane bound via its C-terminal segment. So, to better evaluate the role of the N-terminal segment of RDH11 and the C-terminal segment of RDH8 in the membrane binding of these proteins, different variants (Long and Short) of the aforementioned segments have been studied. In addition, mutations of the C-terminal segment of RDH8 have been introduced to monitor their interaction with lipid monolayers or bilayers.
We have thus analyzed the secondary structure content of these segments by conventional spectroscopic methods such as circular dichroism (CD) and attenuated total reflectance (ATR) infrared spectroscopy whereas their interaction with phospholipids have been mainly monitored by surface pressure measurements when using monolayers and fluorescence spectroscopy for bilayers. Overall, we found that the N-terminal segment of RDH11 adopts an α-helix conformation acting as a transmembrane domain. Values of maximum insertion pressure (MIP) and synergy suggested a preferential binding of the RDH11 Long-peptide to phosphoethanolamine, which are abundant in the RPE. In the case of RDH8, our findings suggest an important role of the long C-terminal segment in membrane binding, which is supported by its helical content and the larger values of MIP and synergy.
We also compared the behavior of RDH8 and its truncated form (RDH8t, without its C-terminal segment) to better understand the involvement of this segment in membrane binding. Thus, both enzymes have been expressed in E. coli, purified by affinity chromatography and studied by the spectroscopic methods mentioned above and by using MIP and synergy measurements. RDH8 and RDH8t display a secondary structure content in agreement with their predicted Rossmann fold.
Interestingly, the removal of the C-terminal segment decreased the temporal and thermal stability of these enzymes. In addition, this segment contributes to protein-lipid interaction especially in presence of negatively charged phosphoprotein-lipids likely through electrostatic interactions. The involvement of the C-terminal segment of the RDH8 in its membrane anchoring has been further confirmed by fluorescence measurements of its two Trp residues located in this segment.
The present characterization of RDH8 is a first step paving the way for the elucidation of its structural and functional features to gain a better understanding of its role within the visual cycle and investigating mechanisms of retinal pathogenesis.
Table des matières
Résumé ... iii
Abstract ... v
Table des matières ... vii
Liste des figures ... xi
Liste des tableaux ... xii
Liste des abréviations ... xiii
Remerciements ... xv Avant-propos ... xvii Chapitre 1 Introduction ... 1 1.1 L’œil ... 1 1.2 La rétine ... 2 1.3 Les photorécepteurs ... 4 1.3.1 Description structurale ... 4 1.3.2 Caractéristiques fonctionnelles ... 5
1.3.3 La biochimie des photorécepteurs ... 6
1.4 L’épithélium pigmentaire rétinien ... 11
1.4.1 Approvisionnement en rétinoïdes ... 12
1.4.2 Recyclage des photorécepteurs ... 13
1.4.3 Absorption de la lumière éparse ... 13
1.4.4 Barrière physique et interface d’échange ... 13
1.5 Le cycle visuel ... 14
1.6 Les RDHs dans le cycle visuel ... 18
1.6.1 Localisation et liaison membranaire des RDHs ... 18
1.6.2 Structure et fonction des RDHs ... 20
1.7 Approches expérimentales de l’étude des interactions entre les protéines/peptides et les membranes ... 26
1.7.1 Les monocouches de Langmuir : modèle d’étude membranaire ... 26
1.7.2 Mesure de pression de surface et détermination de la pression d'insertion maximale ... 28
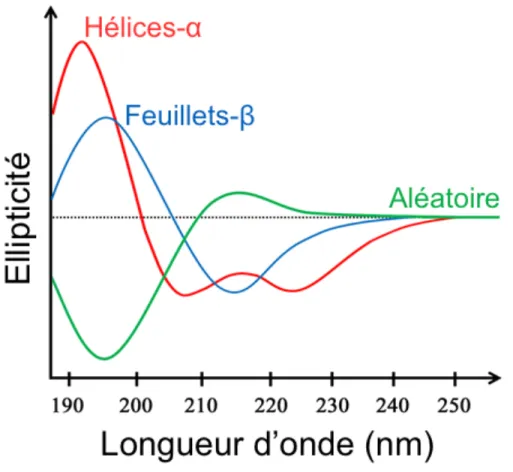
1.7.3 Analyses spectroscopiques de structure secondaire ... 31
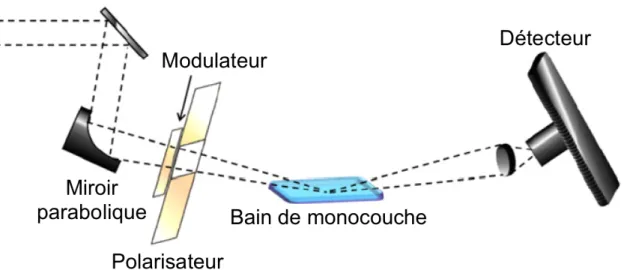
1.7.4 La spectroscopie infrarouge de réflexion-absorption par modulation de polarisation (PM-IRRAS) ... 35
1.7.5 La spectroscopie de fluorescence ... 37
Chapitre 2 Structure of the N-terminal segment of human retinol dehydrogenase 11 and its preferential lipid binding using model membranes
... 43
2.1 Résumé ... 44
2.2 Abstract ... 45
2.3 Introduction ... 46
2.4 Materials and methods ... 47
2.4.1 Materials ... 47
2.4.2 Methods ... 48
2.5 Results ... 50
2.5.1 Analysis of the primary sequence of the Long- and Short-peptides ... 50
2.5.2 Determination of the secondary structure of the Long- and Short-peptides in solution ... 51
2.5.3 Binding of the Long- and Short-peptides to different lipid monolayers ... 53
2.5.4 Secondary structure of the Long- and Short-peptides upon binding to phospholipid monolayers ... 55
2.6 Discussion ... 56
2.6.1 Effect of solvents and of the adsorption of the pure Long- and Short-peptides in monolayer on their structure ... 56
2.6.2 Effect of lipid monolayers on the structure of the Long- and Short-peptides and their preferential binding to phosphoethanolamine ... 57
2.7 Acknowledgements ... 59
2.8 References ... 60
2.9 Tables ... 64
2.10 Figures ... 65
2.11. Supplementary data ... 71
Chapitre 3 Secondary structure and membrane binding of the RDH8 C-terminal segments ... 76
3.1 Résumé ... 77
3.2 Abstract ... 78
3.3 Introduction ... 79
3.4 Materials and methods ... 80
3.4.1 Materials ... 80
3.4.2 Methods ... 81
3.5 Results ... 86
3.5.1 Analysis of the sequence of the Long- and Short-bRDH8 C-terminal segments ... 86
3.5.2 Determination of the secondary structure of the Long- and Short-bRDH8 C-terminal segment ... 86
3.5.3 Binding of the Long- and Short-bRDH8 C-terminal segments to lipid
monolayers ... 88
3.5.4 Binding of the Long- and Short-bRDH8 C-terminal segments to lipid bilayers ... 89
3.5.5 Binding of the acylated Short-bRDH8 C-terminal segments to lipid bilayers ... 90 3.6 Discussion ... 92 3.7 Acknowledgements ... 95 3.8 References ... 96 3.9 Tables ... 99 3.10 Figures ... 100 3.11 Supplementary information ... 107
Chapitre 4 Comparative Study of Retinol Dehydrogenase 8 (RDH8) and its truncated form ... 109
4.1 Résumé ... 110
4.2 Abstract ... 111
4.3 Introduction ... 112
4.4 Material and Methods ... 113
4.4.1 Materials ... 113
4.4.2 Cloning and expression of RDH8 and RDH8t ... 114
4.4.3 Protein sample preparation ... 115
4.4.4 Protein purification ... 115
4.4.5 Electrophoresis and western blot analyses ... 116
4.4.6 Circular dichroism analysis ... 116
4.4.7 Langmuir monolayer study ... 117
4.4.8 Preparation of phospholipid vesicles ... 117
4.4.9 Fluorescence spectroscopy measurements ... 118
4.5 Results ... 118
4.5.1 Purification of recombinant RDH8 and RDH8t ... 118
4.5.2 Assessment of the secondary structure of RDH8 and RDH8t ... 119
4.5.3 Analysis of the temporal and temperature stability of RDH8 and RDH8t ... 120
4.5.4 Determination of the membrane binding of RDH8 and RDH8t ... 121
4.5.5 Evaluation of RDH8 membrane binding using its intrinsic fluorescence ... 123 4.6 Discussion ... 124 4.7 Acknowledgements ... 126 4.8 References ... 127 4.9 Tables ... 132 4.10 Figures ... 133
Chapitre 5 Conclusions et perspectives des travaux de recherche ... 142
5.1 Caractérisation structurale et de liaison membranaire du segment N-terminal de la RDH11 ... 142
5.2 Étude de la liaison membranaire du segment C-terminal de la RDH8 en monocouche et en bicouche lipidique ... 145
5.3 Surexpression, purification et étude comparative entre la RDH8 complète et sa forme tronquée ... 149
Chapitre 6 Bibliographie ... 159
Chapitre 7 Annexes ... 174
Annexe1. Comparison between the behavior of different hydrophobic peptides allowing membrane anchoring of proteins ... 175
Annexe2. Retinol dehydrogenases: membrane-bound enzymes for the visual function ... 244
Liste des figures
Figure 1 : Schéma anatomique de l’œil humain ... 2
Figure 2 : Coupe transversale d’une rétine (A) et représentation schématique de ses différents types cellulaires (B) ... 3
Figure 3 : Schéma simplifié des photorecepteurs de la rétine ... 5
Figure 4 : Structure de l’acide docosahexaénoïque 22:6ω3 (DHA) ... 7
Figure 5 : Schéma simplifié de la rhodopsine ... 8
Figure 6 : Spectres d’absorption des differents pigments visuels ... 10
Figure 7 : Schéma simplifié de l’épithélium pigmentaire rétinien ... 12
Figure 8 : Schéma classique du cycle visuel ... 16
Figure 9 : Localisation des différentes RDHs dans la rétine ... 19
Figure 10 : Alignement de la séquence en acides aminés des différentes RDHs visuelles ... 21
Figure 11 : Modèles conformationnels obtenus à partir des prédictions structurales des différentes RDHs visuelles avec ITASSER (A) et leur superposition selon le Rossman fold (B) ... 24
Figure 12 : Modèles membranaires plans utilisés pour mimer les membranes biologiques et étudier les interactions protéine-lipide ... 27
Figure 13 : Mesure de la pression de surface : représentation simplifiée de l’interaction protéine/peptide – lipide ... 28
Figure 14 : Mesure de la pression de surface : paramètres de liaison ... 30
Figure 15 : Diagramme de la liaison peptidique montrant l’orientation des transitions n ⇒ Π* et Π ⇒ Π* des dipoles exploités en CD ... 31
Figure 16 : Spectres CD caractéristiques des structures secondaires protéiques 32 Figure 17 : Schéma récapitulatif du principe de l’ATR ... 32
Figure 18 : Schéma du montage expérimental d’un PM-IRRAS ... 36
Figure 19 : Diagramme de Jablonski ... 38
Figure 20 : Spectres typiques d’excitation et d’émission des acides aminés aromatiques ... 39
Figure 21 : Acide Fmoc-2,3-diaminopropionique avec une chaîne palmitoyle .... 146
Figure 22 : Exemple d’interaction du segment C-terminal Long-bRDH8-W310R et des vésicules de POPC par la mesure de la fluorescence intrinsèque ... 149
Figure 23 : Spectre infrarouge de la RDH8 (A) et la RDH8t (B) à concentration élevée ... 152
Figure 24 :Détermination de la concentration optimale de la RDH8 et la RDH8t pour les mesures en monocouche ... 153
Figure 25 : Comparaison entre les cinétiques d’adsorption de la RDH8, la RDH8t et du SDS ... 154
Figure 26 : Stabilité de la RDH8 (A) et la RDH8t (B) en fonction du temps : variation du pourcentage en hélice α ... 155
Figure 27 : Chromatogramme préliminaire présentant la mesure par HPLC de la réaction de réduction du tout-trans rétinal (Ral) en tout-trans rétinol (Rol) après l’ajout de lysats cellulaires contenant la RDH8 ... 157
Liste des tableaux
Tableau 1 : Comparaison des caractéristiques des cônes et bâtonnets de la rétine
humaine ... 6
Tableau 2 : Pourcentage d’identité de la séquence en acides aminés entre les
différentes RDHs visuelles humaines ... 21
Tableau 3 : Prédiction de la préférence des RDHs vis-à-vis des cofacteurs ... 22 Tableau 4 : Fréquences typiques de la bande amide I associées aux différentes
structures secondaires en spectroscopie IR ... 35
Tableau 5 : Résumé des différentes essais tentés dans la production de la RDH8
Liste des abréviations
β-OG : n-Octyl-β-D-glucopyranoside µL : microlitres
A2E : N-rétinylidène-N-rétinyléthanolamine
ADN : acide désoxyribonucléique ATR : attenuated total reflection
C12E8 : octaethylene glycol monododecyl ether
CHAPS : 3-[(3-Cholamidopropyl)dimethylammonio]-1-propanesulfonate CHO : chinese hamster ovary
CRALBP : cellular retinaldehyde-binding protein CRBP1 : cellular retinol-binding protein type 1 Ctrl : contrôle
DAPI : 4',6-diamidino-2-phénylindole DDM :n-dodecyl-β-D-maltoside DHA : acide docosahexaénoïque DHRS3 : dehydrogenase/reductase 3 DPH : diphenylhexatriène
E. coli :Escherichia coli
EPR : épithélium pigmentaire rétinien FMN : flavine mononucléotide
His : histidine
HPLC : high pressure liquid chromatography IRBP : interphotoreceptor retinoid-binding protein IRE : élément interne de réflexion
IRRAS : spectroscopie IR de réflexion par absorption LDAO :lauryldimethylamine-N-oxide
LDR : long-chain dehydrogenase/reductase LRAT : lecithin retinol acyltransferase MBP: maltose binding protein
MDR : medium-chain dehydrogenase/reductase Met : méthionine
min : minutes mM: millimolaire mN: millinewton
NAD(P) : nicotinamide adénine dinucléotide (phosphate) oxydé NAD(P)H : nicotinamide adénine dinucléotide (phosphate) réduit PC : phosphocholine
PE : phosphoéthanolamine Phe : phénylalanine
PMIRRAS : spectroscopie de réflexion-absorption par modulation de polarisation en infrarouge
PS : phosphosérine Ral : rétinal
RDH : rétinol déshydrogénase RE : réticulum endoplasmique
RPE65 : protéine de 65 kDa spécifique à l’épithélium pigmentaire rétinien Rol : retinol
ROS : rod outer segment
SDR : short-chain dehydrogenase/reductase SDS :sodium dodecyl sulfate
SEB: segments externes des bâtonnets
SEP: segments externes des photorécepteurs STRA6 : stimulated by retinoic acid 6
Trp : tryptophane Tyr : tyrosine
Remerciements
Je voudrais réserver mes remerciements en premier lieu à Christian Salesse, mon directeur de recherche, qui a supervisé mes travaux de recherche et a guidé les étapes charnières de ce doctorat. Je ne saurais pas le remercier assez pour son dévouement pour la recherche, sa rigueur scientifique et sa disponibilité sans limite. Christian a toujours su faire l’équilibre entre un encadrement efficace et une gestion optimale de l’autonomie de ses étudiants, et je lui en suis reconnaissant. Il va sans dire que Christian est apprécié de tous et est considéré comme un mentor par les professeurs et les étudiants. Compléter cette thèse sous sa direction a donc été une expérience riche et formatrice sur le plan personnel et professionnel.
Je voudrais le remercier en particulier pour m’avoir permis de présenter mes travaux de recherche dans divers congrès nationaux (Alberta, Nouvelle-Écosse, Ontario, etc.) et internationaux (Italie, Brésil). Christian m’a toujours permis d’exprimer mes opinions et mettre en avant mes idées tout en suscitant le sens de l’autocritique et du jugement. À cet égard, Christian est un sage conseiller avec un esprit vif qui inspire le respect et le leadership.
Je dois également mentionner que grâce à ses encouragements et son support, j’ai pu persévérer dans l’accomplissement de cette thèse et l’obtention de bourses et de prix tout au long de mon parcours académique. Je souhaite le remercier en particulier pour m’avoir permis de m’impliquer au sein de la communauté étudiante en représentant mes pairs dans divers conseils d’administration et organisations. Je voudrais également remercier Line Cantin, notre professionnelle de recherche pour son aide dans les différentes expériences et saluer son expertise et sa joie de vivre. Ce fut un réel plaisir d’apprendre sous son aile.
J’aimerais aussi remercier les autres membres de l’équipe, les anciens comme les actuels: Audrey, Sarah B, Élodie, Kim, Sarah R, Marie-Ève, Philippe, Sylvain, ainsi que tous les membres du CUO-recherche que j’ai côtoyés le long de mon cheminement et qui ont été des compagnons de route formidables.
Enfin, je voudrais remercier Mathieu avec qui nous avions commencé au CUO-recherche à la même période et qui a été bien plus qu’un collègue, un ami. Et je ne pourrais pas, bien évidemment passer sous le silence la contribution personnelle et professionnelle d’Habib, pour qui je ne porte qu’une estime sincère et une amitié profonde.
En terminant, je voudrais souligner le soutien de mes parents, ma famille et mes amis. Vos encouragements et vos gestes ont toujours été des remèdes pour franchir les moments difficiles et garder le cap. Merci infiniment!
Avant-propos
Les travaux présentés dans le cadre de ce projet de thèse visent à mettre en lumière la famille des rétinol déshydrogénases (RDHs). Le premier chapitre de cette thèse qui sert d’introduction a été écrit dans le but de situer le lecteur dans un contexte physiologique plus général. Il aborde une description graduelle des éléments visuels : l'œil, la rétine, les photorécepteurs et l’épthélium pigmentaire rétinien (EPR). On y décrit également le cycle visuel et le rôle des RDHs dans ce processus central à la vision. Une section sur l’approche expérimentale utilisée lors de mes travaux de recherche conclut ce chapitre d’introduction. Les chapitres subséquents 2, 3 et 4 se présentent sous forme d’articles scientifiques qui sont déjà publiés ou à soumettre pour publication. Les chapitres 2 et 3 portent sur les segments peptidiques qui permettent la liaison membranaire de la RDH11 et la RDH8, respectivement. Le chapitre 2 intitulé « Structure of the N-terminal segment of human retinol dehydrogenase 11 and its preferential lipid binding using model membranes » a été publié en 2015 dans le journal Biochimica et Biophysica Acta
(BBA) – Biomembranes, alors que le chapitre 3 « Secondary structure and
membrane binding of the RDH8 C-terminal segments » sera soumis prochainement au journal Langmuir.
Concernant l’étude de différentes portions du segment N-terminal de la RDH11 présentée dans le chapitre 2, j’ai réalisé l’ensemble des travaux de recherche. J’ai répété les mesures déjà faites par Mario Méthot, un étudiant précédent qui avait débuté l’étude du segment N-terminal long, et fait de nombreuses mesures additionnelles. J’ai rédigé conjointement ce manuscrit avec Christian Salesse. Pour le chapitre 3 portant sur le segment C-terminal de la RDH8, j’ai accompli la totalité des mesures expérimentales présentées dans ce chapitre et j’ai rédigé le manuscrit conjointement avec Christian Salesse. Les peptides acylés de la RDH8 présentés dans le chapitre 3 ont été produits par François Otis dans le cadre d’une collaboration avec le laboratoire de Normand Voyer du département de chimie à l’Université Laval. Le chapitre 4 relate une approche comparative du comportement structural et de liaison membranaire de la RDH8 et sa forme tronquée RDH8t (sans le segment C-terminal). Le clonage de la RDH8 a été
accompli par Élodie Boisselier avec l’aide de Line Cantin alors que j’ai procédé au clonage de la RDH8t et la surexpression de ces deux protéines, la forme complète et tronquée. On trouve également dans ce dernier chapitre de résultats, le protocole que j’ai mis au point et qui a servi à purifier ces deux protéines. J’ai également exécuté l’ensemble des mesures expérimentales pour caractériser ces deux protéines. J’ai rédigé le manuscrit du chapitre 4 « Comparative study of Retinol Dehydrogenase 8 (RDH8) and its truncated form » conjointement avec Christian Salesse qui sera soumis sous peu au journal Experimental Eye
Research.
La section « Conclusions et perspectives de recherche » constitue le cinquième chapitre de cette thèse. La thèse a été rédigée en français, à l'exception des chapitres 2, 3 et 4 qui se présentent sous forme d’articles scientifiques rédigés en langue anglaise. Les références de ces chapitres sont incluses à la fin de chaque article alors que les références de la thèse (chapitres 1 et 5) sont présentées à la fin du document et constituent le chapitre 6 de « Bibliographie ». Finalement, deux publications dont le contenu est en lien avec cette thèse et dont je suis premier auteur sont jointes en « Annexes ». La première publication annexée, sous forme de revue mais incluant des résultats expérimentaux, est intitulée « Comparison between the behavior of different hydrophobic peptides allowing membrane anchoring of proteins », a été publiée dans le journal Advances in Colloid and
Interface Science en 2014. La deuxième publication annexée constitue également
une revue de la littérature, publiée en 2015 dans Biochemistry and Cell Biology, intitulée « Retinol dehydrogenases: Membrane-bound enzymes for the visual function ».
Chapitre 1 Introduction
1.1 L’œil
L’œil est l’organe responsable de la vision. Il est présent dans une large variété d’espèces plus ou moins complexes: du ver de terre jusqu’à l’Homme. Mais généralement, les yeux des vertébrés sont plus évolués que ceux des invertébrés. Ces derniers sont toutefois seulement capables de distinguer l’intensité et la direction de la lumière.
L’œil humain est formé majoritairement de 3 tuniques [1] : 1) la tunique (externe) fibreuse composée de la cornée transparente en avant et de la sclère opaque en arrière, 2) la tunique (moyenne) uvéale qui est constituée de la choroïde, l’iris et le corps ciliaire, et 3) la tunique (interne) nerveuse essentiellement formée par la rétine. Ces tuniques renferment des milieux transparents essentiels à la transmission de la lumière : l’humeur aqueuse du segment antérieur et le corps vitré (formé de 99% d’eau) du segment postérieur (Figure 1).
L’œil des vertébrés est basé sur un plan structural commun, en comparaison avec l’œil des invertébrés, et selon lequel la lumière traverse la pupille puis le cristallin avant de former une image inversée sur la rétine (Figure 1) [2]. D’ailleurs, l’œil des vertébrés est souvent comparé à une caméra : la pupille de l’iris joue le rôle du diaphragme qui contrôle la quantité de lumière qui pénètre l’œil, le cristallin serait la lentille qui réfracte le faisceau lumineux incident et la rétine correspondrait à une pellicule réutilisable avec les neurones comme « processeurs » pour l’intégration du signal. À la différence d’une caméra, la lentille de l’œil est stationnaire et ce sont les muscles ciliaires qui permettent son accommodation à l’objet, par contraction/dilatation, et vont ainsi modifier son épaisseur et sa courbure pour faire la mise au point de l’image.
L’œil humain adulte est un organe sphérique de 24 mm de diamètre qui pèse environ 7 grammes [3]. Ce petit organe est impliqué dans un tiers de l’information sensorielle transmise au cerveau pour y être traitée. Le cerveau gauche regarde le champ visuel droit et vice versa. Ceci est dû à la disposition spatiale des nerfs optiques qui se projettent du côté opposé du cerveau après s’être croisés au niveau du chiasma optique [1].
Il est intéressant de noter que l’œil humain est sensible à la lumière visible entre 380 et 760 nm [4] et qu’il est capable de distinguer différentes nuances de lumière visible déployant des intensités très contrastées. Cette capacité visuelle accrue est possible grâce à la performance de la rétine.
Figure 1 : Schéma anatomique de l’œil humain (adapté de [5]).
1.2 La rétine
La rétine représente la partie de l’œil excitable par la lumière. Ce tissu qui tapisse le fond de l’œil est commun à tous les vertébrés bien qu’il y ait des variations entre les espèces [6]. La rétine représente l’interface de conversion entre le signal lumineux reçu du champ visuel et le signal électrique acheminé au cerveau (système nerveux central) par le biais du nerf optique pour y projeter une image. La rétine est composée principalement d’une couche externe, l’épithélium pigmentaire rétinien, et une couche interne ou rétine neurale (Figure 2A). Cette dernière est composée de 6 types de cellules neuronales (Figure 2B) : les bâtonnets, les cônes, les cellules bipolaires, les cellules horizontales, les cellules amacrines, et les cellules ganglionnaires [3]. Les cellules de Müller ne sont pas des cellules neuronales mais jouent plutôt un rôle structural de soutien.
La disposition de ces cellules au sein de la rétine respecte une organisation particulière en plusieurs couches (Figure 2A), en fonction des types cellulaires et des organites majoritaires présents dans chaque couche : couche plexifome, synaptique, nucléaire, etc. [2, 7]. Cette organisation présente l’avantage de placer les photorécepteurs à proximité de l’épithélium rétinien pigmentaire (EPR) mais peut être perçue comme contre intuitive dans la mesure où la lumière doit traverser les autres types cellulaires avant d’atteindre les photorécepteurs.
Figure 2 : Coupe transversale de l’organisation cellulaire de la rétine (~ 0.2 mm) (A) et représentation schématique de ses différents types cellulaires (B). EPR : épithélium pigmentaire rétinien, SE : segment externe des photorécepteurs, SI : segment interne des photorécepteurs, CNE : couche nucléaire externe des photorécepteurs, CPE : couche plexiforme externe, CNI : couche nucléaire interne des cellules bipolaires/amacrines/horizontales/Müller, CPI : couche plexiforme interne, CCG : couche des cellules ganglionnaires (adapté de [8, 9]).
1.3 Les photorécepteurs
1.3.1 Description structurale
Les photorécepteurs sont des neurones spécialisés. La rétine est composée de deux types de photorécepteurs qui diffèrent par leur morphologie : les cônes et les bâtonnets (Figure 3). Cette appellation d’ailleurs, tire son origine de la forme de ces cellules. Chaque photorécepteur est constitué de 4 composantes : le segment externe, le segment interne, le noyau et la terminaison synaptique. La lumière traverse les photorécepteurs du côté synaptique vers le segment externe où les photons seront absorbés.
Le segment externe des bâtonnets est composé de compartiments membranaires indépendants de 6 à 8 nm d’épaisseur [10], initialement identifiés par Kühne comme les « plaquettes » [11], maintenant connus sous l’appellation de « disques ». Il y a environ 1000 disques par bâtonnet. Toutefois ce nombre varie selon les espèces [12, 13]. Le recyclage de ces disques se fait du côté apical avec l’intervention de l’EPR où ils seront dégradés (voir section 1.4). Le segment externe des cônes ne contient pas de disques distincts comme les bâtonnets mais il est plutôt formé d’invaginations continues de la membrane plasmique [14]. Les disques membranaires abritent la machinerie enzymatique impliquée dans la phototransduction et le cycle visuel. Le segment interne, relié au segment externe via le cil connecteur, contient les éléments métaboliques des photorécepteurs : mitochondries, golgi, réticulum endoplasmique, cytosquelette, etc. Le noyau se présente quant à lui comme le siège de l’information génétique de ces cellules nerveuses. Enfin, la terminaison synaptique des photorécepteurs comprend un axone qui sert de relais neuronal avec les dendrites des cellules horizontales et bipolaires afin de transmettre l’influx nerveux. Les cellules ganglionnaires prennent ensuite le relais dans la transmission de cet influx vers le cerveau (cortex visuel) par le biais du nerf optique. On parle de sphérule et pédicule pour désigner la terminaison synaptique des bâtonnets et des cônes, respectivement [2].
Figure 3 : Schéma simplifié des photorécepteurs de la rétine [8].
1.3.2 Caractéristiques fonctionnelles
La différence entre les caractéristiques morphologiques des photorécepteurs observée par Schultze en 1866 est à la base de sa « théorie double » sur la fonction visuelle (Duplex retina theory) [15]. Tout d’abord, il est important de noter que seulement 5% des photorécepteurs sont des cônes (∼4.6 millions) principalement localisés au niveau de la fovéa et impliqués dans la vision photopique. Ainsi, 95% des photorécepteurs restants sont des bâtonnets (∼92 millions) responsables de la vision scotopique [16].
Les cônes, saturables à de fortes intensités lumineuses, déploient une réponse rapide et sont responsables de la perception des couleurs [17, 18]. Les cônes sont donc impliqués dans la vision photopique ou la « vision de jour ». Chaque cône est relié à une seule cellule neuronale (ganglionnaire), ce qui confère aux cônes une meilleure acuité visuelle et une résolution spatio-temporelle supérieure. Les bâtonnets sont plus nombreux et déploient une sensibilité supérieure qui leur permet de détecter un photon à la fois. Ils sont notamment responsables de la vision scotopique ou « vision de nuit ». Toutefois, ils sont plus longs à réagir et aisément saturés. Ils déploient également une
latence après une exposition prolongée à la lumière. Chaque bâtonnet est connecté à plusieurs cellules neuronales produisant une vision relativement floue et de faible résolution. Des éléments comparatifs des cônes et bâtonnets sont résumés dans le Tableau1.
Cônes Bâtonnets
Vision Photopique Scotopique
Acuité spatiale Extrêmement élevée Très basse Vitesse de réponse Très rapide
(au-delà de 100 Hz) Lente Gamme de fonctionnement par
rapport à la lumière Énorme Restreinte
Saturation Ne saturent pas en
lumière constante Saturent au crépuscule Adaptation à la lumière Rapide
(loi de Weber sur une large gamme) (à basse intensité, loi quadratique) Lente Sensibilité au contraste Élevée (détecte 0.5%) Basse (5%)
Reprise après
photo-blanchiment Après 20 ms Après 20 min
Adaptation à la noirceur après photo-blanchiment
Sensibilité retrouvée après 5 min
Sensibilité retrouvée après 40 min
Vision en couleur Trichromatique ----
Proportion dans la rétine 5% 95%
Proportion dans la fovéa 100% 0%
Tableau 1 : Comparaison des caractéristiques des cônes et bâtonnets de la rétine humaine (adapté de [19]).
1.3.3 La biochimie des photorécepteurs
Les segments externes des bâtonnets (SEB) sont constitués de 50% de lipides et 50% de protéines (p/p) [20]. Les lipides sont majoritairement des phospholipides, les protéines étant principalement la rhodopsine. Chez les vertébrés, la membrane des
photorécepteurs est très fluide. En effet, la viscosité des disques membranaires est estimée à 2 poises [21]. La fluidité (φ) étant inversement proportionnelle à la viscosité (η), la fluidité des photorécepteurs est comparable à celle de l’huile d’olive. Cette fluidité permet à la rhodopsine d’afficher une diffusion rotationnelle et translationnelle importante [21-23]. Toutefois, cette fluidité est plus limitée chez les invertébrés [24, 25] ou dans les microdomaines lipidiques [26]. La fluidité membranaire est intimement reliée à la composition lipidique. Dans les membranes des segments externes des bâtonnets, il y a principalement 3 types de phospholipides : environ 38% de phosphoéthanolamine (PE), 33% de phosphocholine (PC) et 12% de phosphosérine (PS). On retrouve également environ 10% de cholestérol [20]. Cette composition comporte une distribution asymétrique dans les disques des photorécepteurs entre le feuillet membranaire interne et externe. En effet, le feuillet externe contient majoritairement les phospholipides de type PE (77-87%) et PS (77-88%). Les phospholipides de type PC sont plutôt localisés dans le feuillet interne (65-100%) [27].
La fluidité des membranes des SEB est due principalement à la prépondérance des phospholipides polyinsaturés, estimée à 60% [28]. L’acide docosahexaénoïque ou 22:6ω3 (DHA), schématisé dans la Figure 4, représente à lui seul environ 80% de toutes les chaines acyles polyinsaturées [29]. Le tissu où on retrouve la plus grande concentration corporelle en DHA est la rétine. Il est également intéressant de noter que le degré d’insaturation est aussi réparti de façon hétérogène dans les disques membranaires. Globalement, le feuillet externe contient plus de chaînes acyles insaturées [27]. Cette abondance des phospholipides polyinsaturés rend ces cellules vulnérables vis-à-vis de l’oxydation, encore plus en présence de la lumière qui peut induire la formation d’espèces réactives à l’oxygène [30, 31]. On peut alors comprendre aisément la nécessité de renouveler en permanence les membranes des photorécepteurs (voir section 1.5).
Il existe également plusieurs protéines caractéristiques dans les photorécepteurs, la plus connue étant la rhodopsine, comme mentionné précédemment [32]. Cette protéine représente environ 80-90 % de toutes les protéines discales et occupe environ la moitié de la surface des disques [33-35]. Elle est également présente au niveau de la membrane plasmique [36] mais nous ne connaissons pas à l’heure actuelle les différences fonctionnelles entre ces deux populations de rhodopsine. La rhodopsine bovine, largement étudiée, est constituée d’une partie protéique appelée « opsine » formée de 348 acides aminés et du chromophore 11-cis rétinal attaché à cette opsine via une base de Schiff au niveau de la lysine 296 (Figure 5). L’absorption d’un photon induit la photoisomérisation du 11-cis rétinal en tout-trans rétinal.
Figure 5 : Schéma simplifié de la rhodopsine (adapté de [32]) montrant les 7 domaines transmembranaires hélicaux (cylindres jaunes) reliés par des boucles intra (C-I, II, III) et extracellulaires (E-I, II, III). La Lysine (K296, pourpre) représente le site de liaison du chromophore (non représenté) alors que le glutamate (E113, bleu clair) fournit le contre ion pour la base de Schiff protonée. À noter, la palmitoylation des deux cystéines (C322-323, vert) du côté C-terminal.
La rhodopsine subit plusieurs modifications post-traductionnelles pour être fonctionnelle, comme la glycosylation et la palmitoylation. La rhodopsine appartient à la famille des récepteurs couplés aux protéines G (GPCR) et déploie donc 7 domaines transmembranaires [32]. En plus de son implication centrale dans la phototransduction et le cycle visuel, la rhodopsine a également un rôle dans la formation des disques membranaires. En effet, les souris Knock-Out (KO) pour la rhodopsine n’ont pas de segment externe normal mais seulement une protrusion membranaire [37]. Dans le cas des souris avec un seule allèle, les bâtonnets sont d’apparence normale mais avec moins de rhodopsine [38] alors que pour les souris surexprimant la rhodopsine, les segments externes sont de plus gros diamètre [39].
La majorité des fondements scientifiques concernant les photorécepteurs ont d’abord été établis pour les bâtonnets, car ils sont plus faciles à isoler et plus stables dans le temps (pour une revue, voir [40, 41]). Les pigments visuels des cônes ont donc été relativement moins étudiés. Néanmoins, il est bien établi qu’il existe 3 opsines différentes responsables de la perception des couleurs chez l’Homme : MWS, LWS et SWS1 (Mid, Long et Short-wavelength sensitive, respectivement) qui absorbent dans le vert (535 nm), le rouge (575 nm) et le bleu (445 nm), respectivement (Figure 6). Ces 3 opsines ont été découvertes et identifiées grâce au travail précurseur de Nathans et al. sur les pigments visuels pendant les années 80 [42-44]. En effet, il a été démontré que chez l’Homme, la rhodopsine et le pigment bleu étaient portés sur les chromosomes 3 et 7 alors que les pigments rouge et vert étaient portés conjointement sur le chromosome X [44, 45]. Il est d’ailleurs intéressant de remarquer que ces deux derniers pigments partagent 96% d’identité dans leur structure primaire (seulement 15 acides aminés différents). Malgré une structure tertiaire identique, une analyse poussée de plusieurs mutants a mis en évidence que leur différence spectrale est due à 7 acides aminés essentiels [46]. D’un autre côté, les pigments vert/rouge partagent environ 43-44% d’identité de séquence avec le pigment bleu.
Figure 6 : Spectres d’absorption des differents pigments visuels [8].
La présence de ces 3 pigments visuels chez l’Homme (et autres primates supérieurs) lui permet de bénéficier d’une vision trichromatique [47]. Les poules et les pigeons, bien que moins évolués, profitent d’une vision tétrachromatique grâce à la présence d’un pigment additionnel [48, 49]. D’autres mammifères comme les rongeurs déploient plutôt une vision dichromatique, n’ayant que les pigments SWS avec MWS ou LWS [50]. Certains amphibiens, poissons et reptiles disposent de 5 classes de pigments visuels sur la base des différents types d’opsines : rhodopsine (Rh1), LWS, MWS (Rh2), SWS1, et SWS2 [41, 51, 52]. Toutefois, le nombre de pigments visuels ne corrèle pas toujours avec le degré de vision chromatique et ne traduit pas pour autant un pouvoir spectral discriminatoire. En effet, la crevette-mante possède l’un des systèmes optiques les plus sophistiqués du règne animal avec 12 pigments visuels différents [53]. Ce système visuel multiple ne permet pas une vision dodécachromatique mais semble aider à une meilleure et plus rapide reconnaissance des couleurs. En règle générale, il y a un seul type de pigment par photorécepteur, ce qui détermine le spectre d’absorption et la sensibilité du photorécepteur. Toutefois, il est maintenant établi que les cônes de plusieurs mammifères, tels les souris et les lapins, peuvent contenir conjointement les pigments MWS et SWS [54-56].
1.4 L’épithélium pigmentaire rétinien
L’épithélium pigmentaire rétinien (EPR) représente la barrière sang-rétine qui contrôle les échanges hémato-rétiniens au niveau de l’œil [57]. L’EPR contient environ 3.5 millions de cellules et forme une monocouche de cellules cubiques (~14 µm x 12 µm) et hexagonales (Figure 7A). Les cellules de l’EPR sont incapables de se diviser [58], ce qui accroît les dommages accumulés dans le temps. D’ailleurs, leur densité uniforme au départ (4 000 cellules/mm2) diminue avec l’âge, notamment en périphérie (2 000 cellules/mm2 après 40 ans) [57].
Les cellules de l’EPR sont caractérisées également par leur polarité avec des organites plus présents du côté apical (rétine neurale), tels que les granules pigmentaires alors que d’autres organites sont situés plutôt du côté basal (choroïde) comme le noyau et les mitochondries [59, 60]. Notons également que cette polarité s’applique également aux protéines de l’EPR : les pompes ATPases Na+/K+ se retrouvent du côté apical où il y a également les jonctions serrées et les microvillis [61], alors que les canaux ioniques « bestrophine » sont situés uniquement du côté basal [62].
Généralement, une cellule de l’EPR entre en contact avec 30 à 50 photorécepteurs [63]. L’EPR accomplit plusieurs fonctions spécialisées et essentielles en lien avec cellules avoisinantes dont : 1) fournir aux photorécepteurs les rétinoïdes emmagasinés dans les rétinosomes ou en provenance de la circulation sanguine choroïdale [64], 2) recycler les photorécepteurs par phagocytose [65], 3) absorber la lumière éparse par les granules de mélanine [66], et 4) constituer une barrière physique et une interface d’échange (barrière rétine-sang) [67].
Figure 7 : Schéma simplifié de l’épithélium pigmentaire rétinien (EPR). A) Disposition des cellules de l’EPR par rapport aux autres cellules adjacentes. CH : choroïde, MB : membrane de Bruch, SE : segment externe des photorécepteurs. B) Coupe transversale d’une cellule de l’EPR en contact avec les photorécepteurs. RE : réticulum endoplasmique. Schéma adapté de [68].
1.4.1 Approvisionnement en rétinoïdes
L’EPR est un carrefour pour le métabolisme des rétinoïdes. Ce flux implique la vitamine A issue de la circulation sanguine ainsi que les rétinylesters (stables et non toxiques) emmagasinés dans les rétinosomes et les chromophores provenant du cycle visuel. Ainsi, l’EPR contribue à réguler l’homéostasie des rétinoïdes pour supplémenter
adéquatement les photorécepteurs en 11-cis rétinal, grâce aux différentes enzymes agissant sur ces molécules : STRA6 (stimulated by retinoic acid 6 protein), CRBP1 (retinol binding protein 1), CRALBP (cellular retinaldehyde binding protein), LRAT (lecithin-retinol acyltransferase), RPE65 (retinal pigment epithelium protein 65), RDHs (retinol dehydrogenases), etc. Ce rôle accompli par l’EPR est essentiel pour le bon déroulement du cycle visuel (voir section 1.5).
1.4.2 Recyclage des photorécepteurs
Cette fonction de turn-over a été suggérée initialement par Bairati et Orzalesi en 1963 [69] et démontrée par Young en 1967 par un suivi de la synthèse protéique incorporant la méthionine tritiée [70]. Les photorécepteurs sont exposés à beaucoup de lumière, ce qui induit des dommages au niveau protéique et lipidique [71]. D’où l’importance d’un renouvèlement en continu des photorécepteurs par un processus de phagocytose : environ 1/10 est renouvelé à chaque jour [72]. La quantification des phagosomes internalisés par l’EPR suggère un processus plus rapide pour les bâtonnets que pour les cônes [73].
1.4.3 Absorption de la lumière éparse
La rétine est un milieu propice au stress oxydatif par son exposition à la lumière, sa consommation élevée en oxygène, et la prépondérance de compartiments lipidiques (disques membranaires des photorécepteurs) qui peuvent être peroxydés (pour une revue, voir [74-77]). La rétine subit donc des dommages photochimiques induits par la lumière. En effet, l’énergie photonique peut produire des radicaux libres, un phénomène accentué par la présence de l’oxygène et l’abondance des lipides polyinstaurés. La rétine peut également subir un dommage photomécanique ou photothérmique [71]. L’EPR aide alors à atténuer ces dommages en absorbant l’excédant de la lumière éparse grâce à ses pigments de mélanine et de lipofuscine, mais il s’expose à son tour à un stress oxydatif.
1.4.4 Barrière physique et interface d’échange
Bien que la rétine soit irriguée par une circulation sanguine distincte, les photorécepteurs sont dépourvus de vaisseaux sanguins. Il est bien établi que l’EPR joue le rôle de barrière physique hémato-rétinienne entre la choroïde et les photorécepteurs.
Cette isolation est notamment possible grâce aux jonctions serrées qui lient les cellules de l’EPR entre elles [78, 79]. L’étanchéité de ces jonctions favorise donc le transport transcellulaire par rapport à la voie paracellulaire [80]. En effet, l’échange de nutriments (glucose, acides gras, acide ascorbique, etc.) se fait de l’EPR vers les photorécepteurs alors que l’eau et les déchets font le parcours inverse [81]. Cette interface d’échange est augmentée par la surface des microvillis (Figure 7B). Enfin, l’EPR sécrète (selon sa polarité) des cytokines comme des facteurs de croissance qui sont importants pour le microenvironnement des photorécepteurs et qui participent plus globalement au développement et la survie de la rétine [82].
1.5 Le cycle visuel
Les premières bases du cycle visuel appelé aussi « cycle des rétinoïdes » ont été posées initialement par Wald au début du 20e siècle [83-86], ce qui lui a valu le prix Nobel en 1967. Wald a été en effet le premier à proposer l’existence d’une partie protéique en plus du chromophore au sein de la rhodopsine, à caractériser les composantes moléculaires de la vision et à déterminer un rôle de la vitamine A dans le processus visuel. Comme décrit dans la section 1.3.3, un dérivé de la vitamine A, le 11-cis rétinal est le chromophore des pigments visuels des cônes et des bâtonnets [32, 51, 87]. Le cycle visuel est initié par l’absorption de la lumière par le pigment visuel, ce qui provoque la photoisomérisation du 11-cis rétinal en tout-trans rétinal et la transition de la rhodopsine vers un état excité accompagné d’un changement conformationnel de la protéine. Chez les vertébrés, cet état excité consiste en plusieurs intermédiaires de la rhodopsine : batho, lumi, méta-I et méta-II avec des échelles de temps très courtes (pico, nano, micro ou millisecondes) [88]. La photoisomérisation est encore plus rapide puisqu’elle est estimée à 200 femtosecondes [89, 90]. Le changement conformationnel de la rhodopsine dans son état méta-II lui permet d’activer une protéine G, la transduscine, et d’initier la phototransduction.
Le tout-trans rétinal se détache ensuite de l’opsine avant d’être réduit directement par la RDH8 en tout-trans rétinol (Figure 8) [91-93]. Toutefois, une portion non négligeable du rétinal réagit avec la phosphoéthanolamine (PE) pour former le N-rétinylidène-PE qui est séquestré du côté luménal de la membrane des disques [94]. Il a été démontré que
cette fraction du rétinal est transférée de la lumière discale au cytosol par l’ABCA4 (ATP
Binding cassette subfamily A member 4) [95], un transporteur transmembranaire
rétinien. La base de Schiff entre le PE et le tout-trans rétinal est hydrolysée avant que ce dernier ne soit réduit par la RDH8 en tout-trans rétinol. Celui-ci est alors transporté vers l’EPR grâce à l’IRBP (interphotoreceptor retinoid-binding protein) qui assure le transport dans l’espace intercellulaire [96] et la CRBP1 qui prend le relais au sein de l’EPR [97]. Une fois dans l’EPR, le tout-trans rétinol subit alors une cascade de conversions enzymatiques pour régénérer le 11-cis rétinal, comme indiqué dans la Figure 8. Il est tout d’abord estérifié par la LRAT pour former le tout-trans rétinylester [98, 99], en majorité le tout-trans rétinylpalmitate [100]. Le tout-trans rétinylester est transformé ensuite par la RPE65 en 11-cis rétinol. La RPE65 dispose donc d’une activité i) isomérase de la liaison trans en cis et ii) hydrolase pour rompre la liaison ester [101]. Mentionons ici que la RPE65 a été identifiée comme « la » rétinol isomérase du cycle visuel par trois groupes différents la même année (2005) [102-104]. Le 11-cis rétinol est oxydé ensuite par une RDH de l’EPR (RDH5, RDH10 ou RDH11) pour régénérer le chromophore 11-cis rétinal. Ce dernier est alors pris en charge par la CRALBP [105] puis transféré aux photorécepteurs pour y reconstituer le pigment visuel et conclure le cycle visuel (Figure 8).
Figure 8 : Schéma classique du cycle visuel (adapté de [106]). CRALBP : cellular
retinaldehyde-binding protein, DHRS3 : dehydrogenase/reductase 3, EPR : épithélium
pigmentaire rétinien, IRBP : interphotoreceptor retinoid-binding protein, LRAT : lecithin
retinol acyltransferase, NAD : nicotinamide adénine dinucléotide oxydé, NADPH :
nicotinamide adénine dinucléotide phosphate réduit, RPE65 : protéine de 65 kDa spécifique à l’épithélium pigmentaire rétinien.
Néanmoins, il existe de plus en plus d’évidences qui suggèrent l’existence d’un cycle visuel alternatif au niveau des cônes et qui ferait appel aux cellules de Müller à la place de l’EPR (pour une revue, voir [107-109]). En effet, les cellules de Müller sont capables de convertir le tout-trans rétinol en 11-cis rétinol par le biais de la DES1 (dihydrocéramide désaturase-1) qui jouerait partiellement le rôle de RPE65 [110]. Certains travaux rapportent l’existence d’une RPE65 spécifique aux cellules de Müller [111]. Par contre, les cellules de Müller sont incapables d’oxyder le rétinol. Cette étape se ferait dans les cônes qui peuvent accomplir l’oxydation du 11-cis rétinol en 11-cis rétinal et ainsi régénérer le pigment visuel [112, 113]. À l’inverse, les bâtonnets ne sont pas capables d’accomplir cette oxydation finale [114]. Mais malgré tout, le cycle visuel des cônes n’est pas suffisant et ne peut subvenir seul au maintien de l’acuité visuelle. En effet, après la délétion de la RPE65 ou la LRAT dans l’EPR, les souris RPE65-/- ou LRAT-/- ont démontré une dégénérescence accélérée des cônes [115-119].
Il est important de rappeler que des mutations des différentes protéines liant les rétinoïdes, qu’elles jouent un rôle de transporteurs ou chaperonnes, sont associées à des maladies dégénératives (dystrophies) rétiniennes [120-122] et peuvent être ciblées éventuellement dans des processus thérapeutiques [123]. Dans le même registre, une défectuosité à chaque étape du cycle visuel ou presque peut mener à une dystrophie rétinienne [124] (pour une liste complète des gènes impliqués dans les maladies dégénératives rétiniennes, voir : www.sph.uth.tmc.edu/retnet). Ces dystrophies sont dues soit à la non-régénération du chromophore ou soit à l’accumulation de produits cytotoxiques (adduits phototoxiques) issus du rétinaldéhyde. Trois stratégies peuvent être généralement entreprises pour traiter ces maladies : 1) par thérapie génique afin de remplacer le gène défectueux [125, 126], 2) par supplémentation avec le chromophore déficient ou l’un de ses précurseurs [127-130], 3) par inhibition du cycle visuel pour ralentir la formation de produits cytotoxiques (A2E) [131, 132].
1.6 Les RDHs dans le cycle visuel
1.6.1 Localisation et liaison membranaire des RDHs
Les rétinol déshydrogénases (RDHs) intervenant dans le cycle visuel peuvent être classées en deux groupes (Figure 9) : les enzymes présentes dans les photorécepteurs, comme la RDH8, la RDH12 et la DHRS3, et celles localisées au niveau de l’EPR comme la RDH5, la RDH10 et la RDH11 [106, 133]. Il faut noter qu’il existe également d’autres RDHs telles la RDH13 et la RDH14 dont l’implication en vision reste à démontrer bien qu’elles soient présentes dans les tissus rétiniens [106].
La répartition tissulaire a été démontrée pour toutes ces RDHs avec différentes techniques, notamment par hybridation in situ ou par immunohistochimie [134, 135]. À l’exception de la RDH8, toutes les autres RDHs ont été observées dans des tissus autres que la rétine, comme la prostate, les testicules, le foie, les reins, le pancréas, etc. [106]. D’où l’appellation prRDH parfois utilisée pour désigner spécifiquement la RDH8 : « photoreceptor RDH » [93]. Autre fait intéressant, la RDH8 est la seule RDH dont on a rapporté des acylations au niveau de son segment C-terminal; elles lui serviraient à lier la membrane des disques des segments externes des photorécepteurs [136]. Toutefois, la liaison membranaire de cette enzyme a été observée bien avant son identification [91, 92].
La DHRS3 (dehydrogenase/reductase 3, appelée aussi retSDR1) est l’analogue de la RDH8 dans les cônes puisqu’elle se retrouve exclusivement dans le segment externe de ces cellules [137]. Toutefois, il a été montré par la suite que la DHRS3 était exprimée dans d’autres tissus où elle est attachée au réticulum endoplasmique (RE) ou aux gouttelettes lipidiques [138], ce qui est compatible avec le stockage des rétinoïdes au niveau de ces gouttelettes. De façon similaire, il a été démontré que la RDH10 (37% d’identité avec la DHSR3) se liait aux membranes des gouttelettes lipidiques durant la synthèse des esters acylés [139], confirmant ainsi le couplage entre l’homéostasie des rétinoïdes et le métabolisme lipidique [140, 141]. Au niveau de l’œil, la RDH10 est majoritairement présente dans l’EPR [142], ainsi que dans les cellules de Müller [143]. À la différence de la DHRS3 où seulement son segment N-terminal serait impliqué dans sa liaison membranaire [137, 138], la RDH10 ferait appel à ses segments en N- et C-terminal pour lier les membranes des mitochondries et des gouttelettes lipidiques de
façon dynamique [139]. La RDH10, la LRAT et la CRBP1 forment alors un métabolon à la surface des gouttelettes lipidiques qui permet le maintien de l’homéostasie du rétinol. D’autre part, la RDH12 est également située dans les photorécepteurs mais dans le segment interne; il a été suggéré qu’elle lie les membranes via son segment N-terminal sur la base de sa grande similarité avec la partie globulaire de la RDH11 (73% d’identité) qui baigne dans le cytosol [106]. Cette hypothèse est supportée notamment par l’absence de glycosylations de la RDH12 [144] et par son profil d’ubiquitination [145].
Figure 9 : Localisation des différentes RDHs dans la rétine (modifé de [146]).
Il a été démontré que la RDH11, présente majoritairement dans l’EPR, en plus du segment interne des photorécepteurs [147, 148], se lierait via son segment N-terminal aux membranes des microsomes de l’EPR [149]. Ce segment très hydrophobe adopte une conformation en hélice α pour médier la liaison membranaire de cette enzyme. De
plus, la longueur de la portion N-terminale impliquée dans cette liaison semble affecter le profil d’interaction de ce segment transmembranaire [150].
La RDH5, première RDH identifiée et clonée [151, 152], a été également découverte dans l’EPR. Son mode d’interaction membranaire n’est pas complètement compris dans la mesure où il a été postulé qu’elle lie les membranes via ses segments en N- et C-terminal, dans une configuration luménale, avec une importance relative pour la portion en C-terminal qui baignerait dans le cytosol [153, 154]. Ce segment joue un rôle particulier puisqu’il inclut certaines mutations résultant en la maladie de Fundus
Albipunctatus [155, 156], en plus d’être impliqué dans l’interaction avec les partenaires
de la RDH5 : RPE65 [157, 158], RDH11 [147], RGR [159] et CRALBP [160, 161]. De façon similaire, la RDH10 est capable aussi d’interagir avec la RPE65 et la CRALBP [162]. D’un autre côté, l’équipe de Napoli a relancé le débat quant à la liaison de la RDH5 en démontrant que la forme murine présentait une topologie cytosolique avec un ancrage à la membrane via seulement son segment N-terminal [163, 164].
Il n’en demeure pas moins que cette capacité de liaison membranaire s’avère très utile dans le contexte des RDHs. Premièrement, cela facilite leur interaction avec d’autres partenaires protéiques du cycle visuel liés à la membrane tels que la RPE65 ou la LRAT. Deuxièmement, les substrats de ces RDHs, en l’occurrence les rétinoïdes, sont des molécules très hydrophobes associées aux membranes [165]. Cette interaction membranaire des RDHs les rapproche donc encore plus de leur substrat pour un gain d’efficience réactionnelle.
1.6.2 Structure et fonction des RDHs
Les RDHs appartiennent à la famille des Short-chain dehydogenases/reductases (SDRs) [166]. Cette famille protéique identifiée pour la première fois au début des années 80 [167] compte au-delà de 163 000 membres qui peuvent être regroupés dans 464 familles [168]. Chez l’Homme, il existe pas moins de 80 gènes codant pour ces enzymes impliquées dans un large spectre métabolique [169]. Les SDRs sont généralement composées de 250 à 350 acides aminés partageant au moins 20-30% d’identité de séquence. Les RDHs visuelles respectent cette règle comme indiqué dans la Figure 10 et le Tableau 2.
Tableau 2 : Pourcentage d’identité de la séquence en acides aminés entre les différentes RDHs visuelles humaines.
Figure 10 : Alignement de la séquence en acides aminés des différentes RDHs visuelles [106]. Les résidus identiques entre toutes les RDHs sont indiqués en rouge et les résidus similaires en jaune. Les 4 étoiles (*) font référence à la tetrade catalytique (N-S-Y-K) alors que le motif riche en glycines (GxxxGxxG) est montré en haut de la figure.
GxxxGxxG
*
* *
*
% RDH5 RDH8 DHRS3 RDH10 RDH11 RDH12 RDH13 RDH14 RDH5 100 35 32 28 29 31 29 28 RDH8 100 29 29 27 27 28 30 DHRS3 100 37 23 24 26 27 RDH10 100 28 24 25 25 RDH11 100 73 51 48 RDH12 100 51 46 RDH13 100 51 RDH14 100On peut en effet remarquer entre autres, la conservation de deux séquences signatures caractéristiques des SDRs :
- Le motif riche en glycines (GxxxGxxG) situé dans le domaine de liaison du cofacteur [170]. La présence d’un acide aminé acide (v.g. acide aspartique ou glutamique) à proximité de ce motif favorise la liaison du NAD grâce à une liaison hydrogène entre le groupement hydroxyle du ribose du cofacteur et l’hydrogène de l’acide aminé en question [170, 171]. Inversement, un acide aminé basique situé dans ce domaine induit une préférence pour le NADP par une stabilisation électrostatique [171, 172]. L’affinité des RDHs vis-à-vis de leur cofacteur peut être prédite in silico [106, 173] comme cela est présenté dans le Tableau 3.
- La tétrade catalytique (N-S-Y-K) formée par les 4 acides aminés impliqués dans la réaction enzymatique (voir *, Figure 10) [174]. La tyrosine (Y) est essentielle car elle joue le rôle d’un catalyseur acide/base pour le transfert d’un hydrure entre le cofacteur et le substrat [166]. La sérine (S) stabilise le substrat tandis que la lysine (K) forme une liaison hydrogène avec le ribose du cofacteur et abaisse le pKa de la tyrosine catalytique pour favoriser la réaction à pH neutre [133, 175]. À l’état inactif, une molécule d’eau mime la présence des groupements OH du substrat et du cofacteur en établissant des liens hydrogène avec la tyrosine et la lysine.
Tableau 3 : Prédiction de la préférence des RDHs vis-à-vis des cofacteurs. Elle a été effectuée avec l’algorithme Cofactory [173]. Le score du cofacteur préféré pour chaque enzyme est montré en gras (plus haut score).
Bien que la préférence pour le cofacteur (ou le sens réactionnel) peut varier d’une RDH à une autre, leur capacité commune de lier un cofacteur découle de la présence d’un motif structural caractéristique nommé le Rossman fold (Figure 11A) [176, 177]. Ce motif est partagé entre toutes les autres SDRs et même les MDRs (Medium-chain
dehydogenases/reductases) ainsi que les LDRs (Long-chain dehydogenases/ reductases) [169, 178]. Le Rossman fold est constitué généralement de 6-7 feuillets β
parallèles entourés de 3-4 hélices α (Figure 11B) [176, 177]. La structure tertiaire de plusieurs SDRs a été résolue démontrant la présence effective du Rossman fold [175]. À ce jour, chez l’homme, une trentaine de structures de SDRs existe dans la banque PDB de structures de protéines (http://www.rcsb.org/pdb). Toutefois, aucune structure n’a été déposée encore pour un membre de la famille des RDHs. Une prédiction structurale basée sur la séquence primaire des RDHs visuelles démontre clairement la présence du motif Rossman fold [106], comme présenté dans la Figure 11A. La prédiction de ce motif structural commun entre les différentes RDHs est supportée par la similarité entre leur séquence et leur fonction et celles des SDRs.
L’activité enzymatique de plusieurs RDHs a été caractérisée in vitro avec différents substrats et cofacteurs [106]. Dans la majorité des cas étudiés, l’analyse a été entreprise avec des homogénats cellulaires où les RDHs étaient capables souvent d’accomplir l’oxydoréduction bidirectionnelle des rétinoïdes fournis [147]. Toutefois, cette approche n’est pas parfaite et ne traduit pas toujours ce qui peut se passer dans le milieu physiologique. Premièrement, il est extrêmement ardu de contrôler les différents éléments chimiques et protéiques présents dans les cellules utilisées pour leur surexpression et qui peuvent avoir un impact positif/négatif sur l’activité enzymatique mesurée. Deuxièmement, les concentrations physiologiques en cofacteur sont difficiles à reproduire avec exactitude (type cellulaire, espèce, etc.), et nul ne peut exclure la présence de rétinoïdes endogènes dans la phase lipidique des membranes de ces extraits cellulaires. Troisièmement, il est plus ardu de quantifier les RDHs dans de telles conditions.
Figure 11 : Modèles conformationnels obtenus à partir des prédictions structurales des différentes RDHs visuelles avec ITASSER (A) et leur superposition selon le Rossman
![Figure 1 : Schéma anatomique de l’œil humain (adapté de [5]).](https://thumb-eu.123doks.com/thumbv2/123doknet/5534488.132319/20.918.204.663.241.599/figure-schéma-anatomique-l-œil-humain-adapté.webp)
![Tableau 1 : Comparaison des caractéristiques des cônes et bâtonnets de la rétine humaine (adapté de [19])](https://thumb-eu.123doks.com/thumbv2/123doknet/5534488.132319/24.918.113.793.276.856/tableau-comparaison-caractéristiques-cônes-bâtonnets-rétine-humaine-adapté.webp)
![Figure 5 : Schéma simplifié de la rhodopsine (adapté de [32]) montrant les 7 domaines transmembranaires hélicaux (cylindres jaunes) reliés par des boucles intra (C-I, II, III) et extracellulaires (E-I, II, III)](https://thumb-eu.123doks.com/thumbv2/123doknet/5534488.132319/26.918.200.676.430.830/simplifié-rhodopsine-montrant-domaines-transmembranaires-hélicaux-cylindres-extracellulaires.webp)

![Figure 11 : Modèles conformationnels obtenus à partir des prédictions structurales des différentes RDHs visuelles avec ITASSER (A) et leur superposition selon le Rossman fold (B) (adapté de [106])](https://thumb-eu.123doks.com/thumbv2/123doknet/5534488.132319/42.918.105.777.118.746/modèles-conformationnels-prédictions-structurales-visuelles-itasser-superposition-rossman.webp)

![Figure 17 : Schéma récapitulatif du principe de l’ATR ([219]). Le faisceau IR se propage dans le cristal trapézoïdal (IRE) par de multiples réfections selon un angle d’incidence θ avec la normale au plan du cristal](https://thumb-eu.123doks.com/thumbv2/123doknet/5534488.132319/52.918.252.651.384.666/schéma-récapitulatif-principe-faisceau-trapézoïdal-multiples-réfections-incidence.webp)