Régulation du complexe suppresseur de tumeurs BAP1/ASXL2 par ubiquitination
93
0
0
Texte intégral
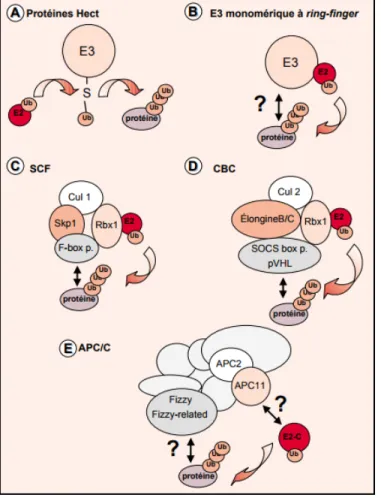
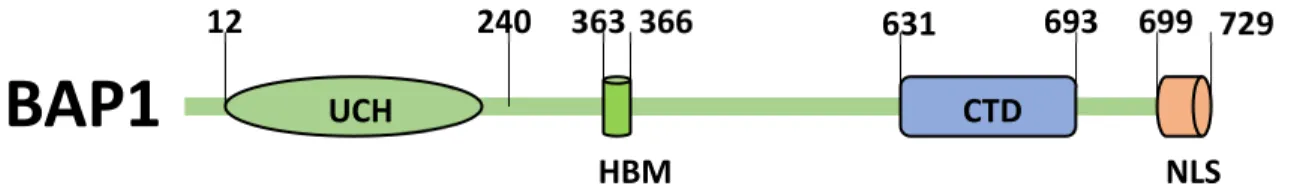
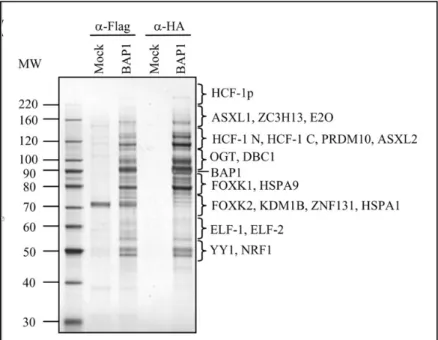
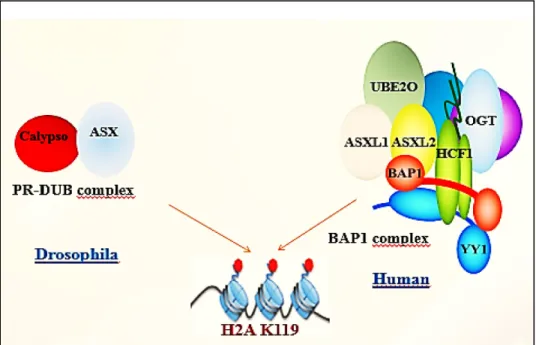
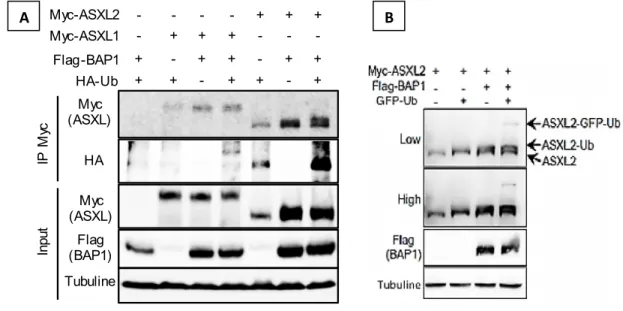
Figure


+7



Outline
Documents relatifs