Relations entre protéolyse hépatique et qualités technologiques et sensorielles du foie gras de canard
202
0
0
Texte intégral
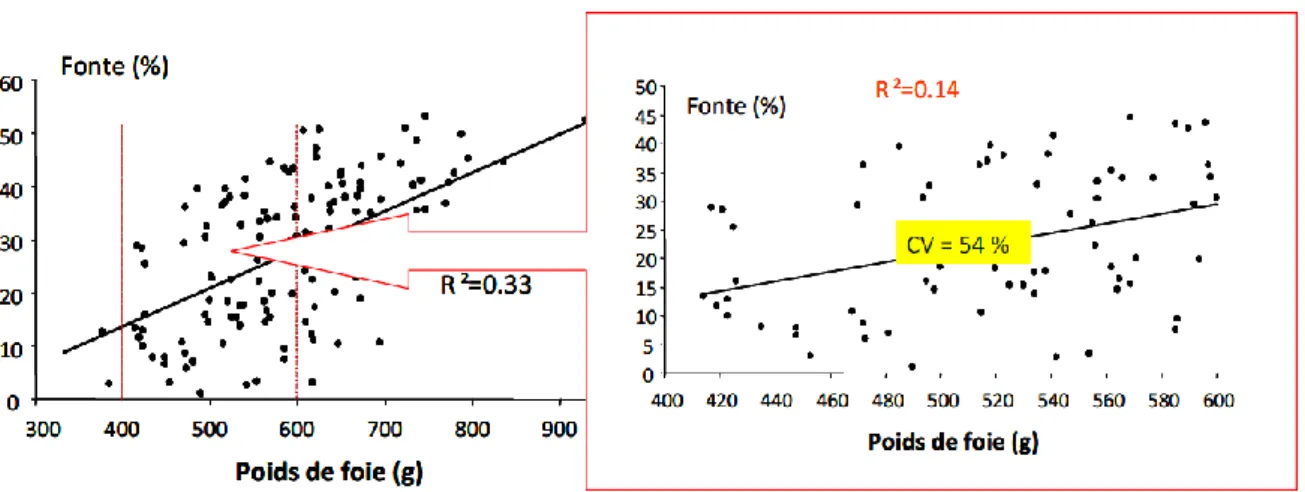
Figure

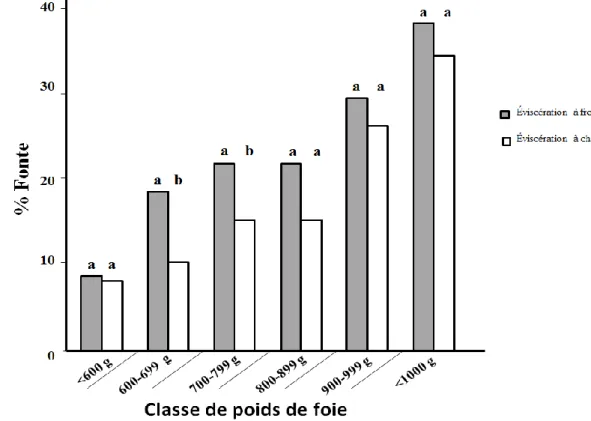
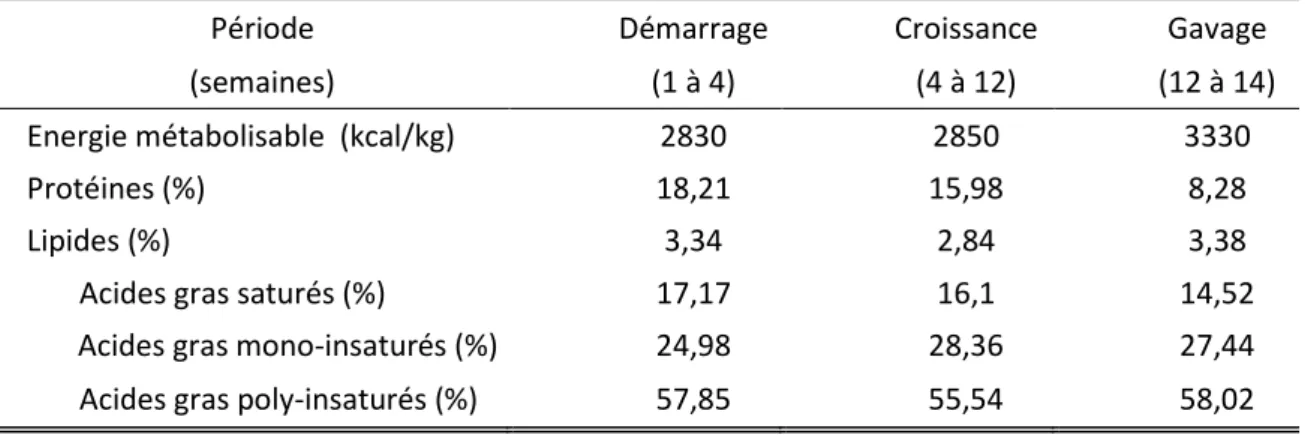
+7

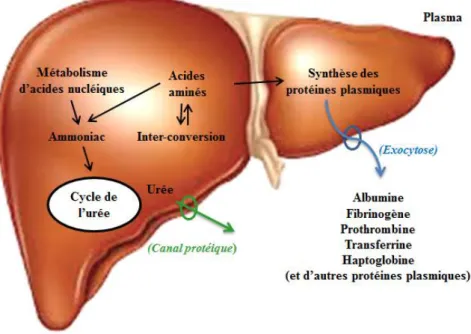



Documents relatifs