L
/
am
é
lioration
d
e
l
a
r
és
i
s
t
ance
d
es
pl
a
nt
es c
ultiv
ées aux
p
aras
i
tes
c
on
s
tim
e
un
o
b
jec
ti
f
m
a
j
e
ur d
e
l
a
plup
ar
t d
es
pro
g
ramm
es
de
sé
l
e
ction
gé
n
é
tique. Au cour
s
du
xxe
s
i
è
cl
e,
un
g
r
an
d nomb
re
d
e v
ari
é
t
és
am
é
-lior
ées o
nr
é
t
é
cr
éées e
t
o
nt p
e
rmi
s
un
e
a
ugm
e
nt
a
ti
o
n d
e
l
a
producti
v
it
é
tout
e
n
limitant
l
'
utilisation
de produit
s
p
es
ti-c
id
es
et fongi
c
ide
s
;
e
ll
e
s r
e
p
rése
nt
e
nt un
e
nj
e
u
co
n
s
id
é
r
a
bl
e
p
o
ur l
'ag
ricultur
e
d
e
d
e
main. Un
e
bonn
e co
nn
aiss
an
ce e
t un
e
c
on
se
rv
a
ti
o
n
a
d
é
qu
a
t
e
d
es
r
ess
our
ces
gé
n
é
riqu
es, ass
o
c
i
ées à
un
e ges
tion r
a
i-s
onn
ée
d
es
g
è
n
es
d
e
r
é
si
s
t
a
n
ce
id
e
ntifi
és,
a
pp
a
r
a
i
ss
ent comm
e
d
es é
l
é
m
e
nt
s
cl
efs
d
a
n
s
la
pour
s
uite d
e ce
tte am
é
li
ora
ti
o
n
.
Au
c
ours d
es
d
e
rni
è
r
es a
nn
ées,
n
o
tr
e
c
onnai
ss
an
ce
d
es s
tr
a
t
ég
i
es
d
e
d
é
f
e
n
se
d
es
plant
es e
t
,
e
n p
a
rti
c
uli
e
r
,
d
es gè
n
es
d
e
r
és
i
s
t
a
n
ce
qui l
es g
ou
ve
rn
e
nt
s'es
t
c
on
s
i
-d
é
r
a
bl
e
m
e
nt
ac
cru
e
.
L
'
obj
ec
tif d
e ce
tte r
e
vu
e es
t d
e
fair
e
un
e
sy
nth
èse
d
e
l
'é
t
a
t
ac
mel d
es c
onn
a
i
s
-s
an
ces c
on
ce
rnant
l
'o
r
ga
ni
sa
ti
on e
t l
'évo
-lution d
es gè
n
es
d
e rés
i
s
t
a
n
ce s
p
éc
ifiqu
e
(R). C
e
t
te sy
nth
èse se
limi
te a
ux
as
p
ec
t
s
e
n rel
a
tion dir
ec
t
e avec
l
a c
on
serva
tion
d
es gè
n
es
R
e
t
l
e
ur
v
al
o
ri
sa
ti
o
n
e
n am
é
-lior
a
ti
o
n d
es
pl
an
te
s
. L
es
fon
c
ti
o
n
s
d
es
gè
n
es
R e
t
,
plu
s gé
n
é
ral
e
m
e
nt
,
l
es
m
éca
-nism
es
d
e
d
é
f
e
n
se
d
es
plan
tes
n
e sero
nt
S. Noir, P. Lashermes: IRD, Laboratoire Geneîrop, BP 5045, F-34032 Montpellier, France.
<[email protected]> Tirés à part: S. Noir
essources génétiques
Organisation et évolution des gènes
de résistance chez les plantes
Sandra
Noir,
Philippe
Lashermes
gu
evo
qu
és,
t
a
ndi
s
qu
e
l
es
po
ss
ibili
tés
d
'a
m
é
li
o
r
a
tion d
e
l
a
r
és
i
s
t
a
n
ce
d
es
plan
tes a
ux mal
a
di
es
p
a
r d
es s
tr
a
t
ég
i
es
d
e
t
y
p
e
biot
ec
hn
o
l
og
igu
e
n
e se
ront p
as
a
bo
r
d
ées
.
Interactions
plantes-parasites ·
la résistance
spécifique de type
gène-à-gène
L
es
pl
a
nt
es
ont d
évelo
pp
é a
u
co
ur
s
d
e
l
e
ur
évo
lution d
e
n
o
mbr
e
u
ses
d
éfe
n
ses
Tableau 1
co
ntr
e
l
es age
n
ts
p
a
th
ogè
n
es [
l
]
. On
p
eu
t
c
i
re
r n
o
t
a
mm
e
nt
:
-
l
'ac
qui
s
iti
o
n d
e ca
r
ac
t
é
ri
s
tique
s
suu
c
tu-r
ales
gui p
e
u
ve
nt
ag
ir
co
mm
e
de
s
b
ar
ri
è
r
es
ph
ys
iqu
es e
t ral
e
nri
r
l
a
pro
g
r
ess
i
o
n d
e
l
'age
nt p
a
tho
gè
n
e a
u
se
in d
e
l
a
plan
te, et
:
-
la p
ro
du
c
tion d
e s
ub
sta
n
ces
to
x
iqu
es
p
o
ur
l'age
nt p
a
tho
gè
n
e
.
S
i
ce
rt
ai
n
s
m
écan
i
s
m
es co
n
st
ituti
fs c
h
ez
l
a
pl
ante so
nt
assoc
i
és à
un
e
d
é
f
e
n
se
pas
s
i-ve [2
], d
'a
utr
es,
induit
s,
p
a
rti
c
ip
e
nt
à
l
'ex
p
ress
i
o
n d
'
un
e
r
és
i
s
tan
ce
lo
c
al
e (
r
é
ac-tion d
'
h
y
p
e
r
se
n
s
ibilit
é,
HR
)
ou
sys
t
é
mique
(
r
és
i
s
tan
ce sys
t
é
miqu
e ac
qui
se o
u indui
re,
SAR/I
S
R
). Ces
m
écani
sm
es
induit
s
limi-tent l
a
di
ssé
min
a
tion d
e
l'
age
nt p
a
th
ogè
ne
a
u
s
ire d
'
in
fec
tion
e
t pr
é
mwlÎ
sse
nt
l
a
plan-te contr
e
un
e agress
ion ult
é
ri
e
ur
e [3, 4].
D
ans
l
e ca
dr
e
d
e ce
tt
e
d
é
f
e
n
se ac
ti
ve,
l
a
rés
i
s
t
a
n
ce
d
e
t
y
p
e gè
n
e-à
-
gè
n
e [5
]
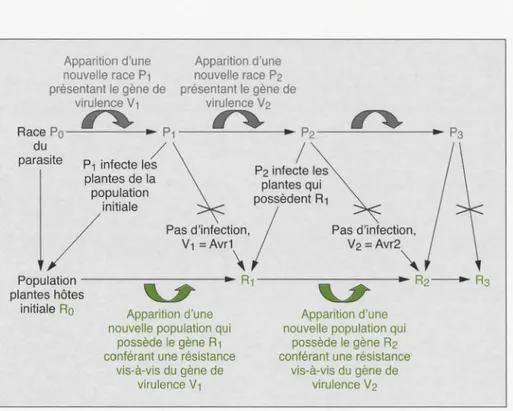
Types d'interactions entre agent pathogène et plante hôte dans le cadre
de la résistance gène-à-gène
Génotype de l'agent pathogène 1
Avr 1, avr 2 avr 1, Avr 2
Génotype de la plante hôte 2
R1 r2 r1 R2 Nature de l'interaction Incompatible Compatible Compatible Incompatible 1 Gène d'avirulence (Avr), gène de virulence (avr). 2 Gènes de résistance fonctionnel (R). non fonctionnel (r).
(tableau 1)
o
cc
up
e
un
e
pl
ace
tr
ès
impor-t
a
nt
e.
Dan
s
l
e cas
d
'
un
e
int
e
ra
c
tion
incomp
a
tibl
e,
la
plant
e
d
é
t
ec
te
l
a
pr
ése
n-ce
d
e
l
'age
nt p
a
tho
gè
n
e e
t p
arv
i
e
nt
à
l
e
n
e
utrali
se
r
:
l
a
pl
a
nt
e es
t di t
e
r
és
i
s
t
a
n
ce e
t
l'
age
nt p
a
tho
gè
n
e es
t
c
on
s
id
é
r
é c
orn m
e
av
irul
e
n
t.
Al
te
rn
a
ti
ve
m
e
nt
,
d
a
n
s
l
e cas
d
'
int
erac
ti
o
n
s co
mp
a
tibl
es,
l
'age
nt p
a
th
o
-gè
n
e
par
v
i
e
nt
à co
loni
se
r l
a
plant
e
h
ôte
et
l
a
mal
a
di
e se
d
év
el
o
pp
e
: l
a
pl
a
nt
e es
t
dit
e se
n
s
ibl
e e
t l
'
age
nt p
a
th
ogè
n
e v
iru
-l
e
nt. La r
és
i
s
t
a
n
ce es
t d
é
t
e
rmin
ée
par
l'int
era
cti
o
n
e
ntr
e
l
e
produit d
'
un
gè
n
e
d
e
l
a
pl
a
nt
e
h
ô
t
e,
l
e gè
n
e
R,
e
t l
e
p
ro
duit
d
'
un
gè
n
e
d
e
l'
age
nt p
a
tho
gè
n
e,
l
e gè
n
e
d
'av
irul
e
n
ce (gè
n
e
Avr).
L
a s
p
éc
i
fic
it
é
o
b
se
r
vé
e
l
o
r
s
d
e
l
'
in
terac
ti
o
n in
com
p
a-tibl
e se
r
a
it
li
ée à
un
évé
n
e
m
e
n
t
pr
écoce
d
a
ns
l
a rés
i
s
t
a
n
ce,
l
'é
t
a
p
e
d
e
r
eco
nn
ais
-sa
n
ce, e
ntr
e
l
es
p
ro
duit
s
d
es gè
n
es
R
e
t
Avr. U
n mod
è
l
e
«é
li
c
it
e
ur
/réce
p
ce
ur
»a
é
t
é
p
ro
p
osé
p
o
ur
ex
pliqu
e
r l'int
e
r
ac
ti
o
n
gè
n
e-à-gè
n
e [6
]. D
ans ce
m
odè
l
e,
l
e gè
n
e
Avr serai
t r
es
p
o
n
sa
bl
e
d
e
l
a
p
ro
du
c
ti
o
n
d
'
un
é
li
c
i
te
ur
(
m
o
l
éc
ul
e
produit
e
p
a
r l
e
p
a
r
as
it
e e
t
reco
nnu
e par
l
a
pl
a
nt
e at
t
a
-qu
ée)
qui
int
e
r
ag
ir
ai
t
avec
un r
éce
p
te
ur
s
p
éc
i
fi
qu
e,
l
e
p
ro
duit du
gè
n
e
d
e
r
és
i
s
-t
a
n
ce. La for
m
a
t
io
n d
e ce co
mpl
exe
indui
rai
t un
s
i
g
n
al
d
o
nt l
a
tr
ans
du
ct
i
o
n
co
nduirai
t à
l
a
mi
se e
n pl
ace
d
'
un
e cas
-ca
d
e
d
'
évé
n
eme
nt
s con
dui
sa
nt
à
l
a rés
i
s
-ran
ce [7]
.
L
a
l
oc
ali
sa
tion d
e
l
a
plu
pa
rt d
es age
nt
s
ph
yto
p
a
th
ogè
n
es po
ur
ra
i
t
l
a
i
sser s
upp
o
-se
r
à to
rt qu
e
l
es
p
ro
t
é
in
es
R
cor
r
es
p
o
n
-d
e
nt
à
d
es
produit
s ex
tr
ace
llul
a
ir
es
d
e
ry
p
e
r
éce
pt
e
ur
s
.
E
n
effet, s
i
ce
rt
a
in
es
p
ro
t
é
in
es
R
(
p
ar e
x
e
mpl
e,
p
ro
t
é
in
es
Xa2
l
e
t
Cf) so
nt
e
ff
ec
ti
ve
m
e
nt
ex
rr
ac
el-lul
a
ir
es,
l
a
m
a
j
o
ri
té
d
es
p
ro
t
éines co
d
ées
p
a
r l
es gè
n
es
R
é
cudi
és sem
bl
e
nt
ê
tr
e
intr
ace
llul
a
ir
es.
D
es sys
t
è
m
es
d
e sécré
-ti
o
n
(
dit
s
d
e ry
p
e
III
) au sein
d
e
b
acté
-ri
es
ph
yco
p
at
h
ogè
n
es o
nt
é
t
é
d
éco
u-ve
rt
s.
Il
s
p
e
rm
e
tt
ra
i
e
nt
l'i
nj
ect
i
o
n d
e
p
ro
t
é
in
es Av
r
a
u
se
in
mê
m
e
d
e
l
a ce
llu
-l
e végé
t
a
l
e
,
offra
n
t
l
a
p
oss
ibilit
é
d
'
un
e
int
e
r
ac
ti
o
n di
rec
t
e et
int
race
llul
a
ir
e
ent
r
e
l
es
p
ro
du
its
R
et Av
r
[8]. Une
ce
ll
e
i
nte
r
ac
ti
on a été
d
é
m
ontrée ent
r
e
le
s
p
ro
duit
s
Pro
e
t
Av
rP
to i
nt
erve
n
a
nt
l
o
r
s
d
e
l
a
r
éac
ti
o
n
i
n
co
mp
a
tibl
e
toma
t
e/
Pseudomonas syringae
p
v
tomato
[9,
1 O
]. Ce
p
e
nd
a
nt
,
p
o
u
r
l
a
plu
part
d
es
p
roté
in
es
R
, a
u
c
un
e
int
erac
t
io
n dir
ecte
avec
l
a
p
ro
t
é
i
ne Av
r n
'a été m
i
se e
n
év
i
-d
e
n
ce.
R
éce
mm
ent,
l
'
exis
t
ence
d
e
p
ro-t
é
in
es c
ompl
é
m
enta
i
res (par exe
mpl
e,
Avr-
r
éce
pt
e
u
r p
u
tatif) ass
ur
a
nt un li
e
n
ph
ys
iq
ue e
nt
re
l
es
p
ro
t
é
in
es
d
es gè
n
es
R
e
t
Avr a é
t
é s
u
ggé
r
ée
[ 11
].
Gènes
de résistance
A
u
co
u
rs
d
es
d
e
rni
è
r
es a
nn
ées,
l
e
cl
o
n
age
d
e
plu
s
i
e
u
rs gè
n
es
d
e rés
i
s
t
a
n
ce e
t l
a
caracté
ri
sa
ti
o
n d
es
p
ro
dui
ts
d
é
duit
s
d
e
l
e
ur
sé
qu
e
n
c
e r
es
p
ec
ti
ve o
ne p
er
mi
s
d
es
ava
n
cées cons
id
é
r
a
bl
es
d
a
n
s
l
a co
nn
ais
-sa
n
ce
d
es
b
ases
m
o
l
éc
ul
aires
d
e
l
a
r
és
i
s
-ta
n
ce s
p
éc
i
fi
qu
e
d
es
pl
a
nt
es.
Ces gè
n
es
R
o
ne
é
t
é
i
s
ol
és
d
'
es
p
èces végé
-tal
es var
i
ées (to
m
ate
,
lin
,
ri
z
,
ta
b
ac, a
r
a
-b
e
rc
e,
b
e
t
te
r
ave s
u
c
ri
è
r
e) [
1
2]
.
M
al
g
r
é
l
'
i
mp
orta
nt
e
di
ve
r
s
it
é de
s p
a
r
as
i
ces
aux-qu
els
il
s
co
n
fè
r
e
nt
l
a
r
és
i
sta
n
ce (c
h
am
pi
-g
n
o
n
s,
b
ac
t
é
ri
es, v
iru
s o
u
e
n
coce
n
é
m
a-to
d
es),
l
e
ur
c
omp
a
r
a
i
so
n
révè
l
e
un
e fo
rt
e
h
omo
l
og
i
e de séq
u
ences a
in
s
i
q
u
e
l
a
co
n
serva
ti
on
d'un
certa
in n
o
mbr
e
d
e
m
ot
i
fs s
tru
c
tur
aux.
Fonctions putatives
des produits de gène
R
P
o
ur
l
a
m
a
j
o
ri
té des gè
n
es
R i
so
l
és,
l
a
pro
t
é
in
e dé
du
ite
d
e
l
e
ur
sé
qu
e
n
ce
nu
cléot
idi
q
u
e parcage p
lu
s
i
e
ur
s é
l
é
m
e
nt
s
s
tru
cturaux avec
d
es
pr
o
t
é
in
es ca
r
ac
t
é
ri
-sées c
h
e
z
l
a
l
ev
u
re,
l
a
d
rosop
hil
e o
u
encore
l
es ve
rt
é
br
és [
1
3]. La
plu
part
d
es
p
ro
du
i
t
s
R
co
mbin
era
i
e
nt un d
o
m
a
in
e
r
éce
p
teur e
t u
n
d
o
m
a
in
e effecte
ur [1
2],
e
t
ass
ur
eraie
nt d
e
u
x fo
n
c
ti
o
n
s maje
ur
es :
l
a
r
ec
onn
a
i
ssa
n
ce
d
e
m
o
l
éc
ul
es é
li
c
i
t
ri
ces
..
LZITIA1
c:J:,
NBS=
=
otif....
~....
~ hydrophobe TPlo LAA RPS2 N 12 RPM1 RPP5 Prl L6 Mi-
Cf9Cahiers Agricultures 2000
;
9
:
301
-
9
p
a
r
des
m
éca
ni
s
m
es
d
'
i
nt
e
r
act
i
o
n
s
p
ro
t
é
i
-n
e-pro
t
é
in
e e
t l'
ac
ti
va
ti
on d
i
rec
t
e o
u
indir
ec
t
e
d
e s
i
g
n
aux
d
e trans
du
ct
i
on
[
7]
.
C
i
n
q p
r
in
c
ip
a
u
x
d
o
m
a
in
es s
tru
c
tur
a
u
x
co
n
servés o
nt
é
t
é
di
s
tin
g
u
és.
L
a
figure
1
illu
s
tr
e ces
di
ffé
r
e
nt
s
d
o
m
a
in
es e
t
l
e
ur
l
oc
ali
sa
ti
o
n.
La
plup
a
rt d
es gè
n
es
R
cl
o-n
és à ce
j
o
u
r p
r
ése
nt
e
nt un
do
m
a
in
e
L
RR
(Leucine-
R
ich
Repeats)
.
Les
d
o
m
a
in
es L
RR
co
rr
es
p
o
nd
e
nt
à
l
a
r
é
p
é
ti
-t
i
on
d
'
un
mo
ti
f
d
e ta
ill
e varia
bl
e
com-pr
e
n
an
t d
es
l
e
u
c
in
es (e
t
/o
u d
'
a
u
t
r
es rés
i
-du
s
h
y
d
ro
ph
o
b
es)
. Il
s
in
te
r
v
i
e
nd
ra
i
e
n
t
d
a
n
s
l
es
m
éca
ni
smes
d
'
i
nterac
ti
ons
p
ro-t
é
in
e-
p
ro
t
é
in
e. Le
d
o
m
a
in
e sé
rin
e/
thr
éo-n
i
n
e k
in
ase se
pr
ése
nt
e se
ul
(
p
a
r
exe
m
p
l
e, pro
dui
t
du
gè
n
e
Pto) o
u bi
e
n
jo
int
à
un d
o
m
a
in
e L
RR
(pa
r
exe
mpl
e
,
p
ro
duit du
gè
n
e
Xa2
1
).
Il
se
r
a
it impli
q
u
é
d
a
n
s
d
es réac
ti
o
n
s de p
h
osp
h
o
r
y
l
ation
assoc
i
ées aux casca
d
es
d
e s
i
g
n
al
i
sat
i
o
n.
L
e
d
o
m
a
in
e N
B
S
(Nucfeotide Binding
Site
)
co
r
res
p
o
nd
à
un
s
i
te
d
e
fi
xa
ti
o
n
et
d
'
h
ydro
l
yse
d
es
nucl
éo
t
i
d
es rr
iph
os
ph
aces
AT
P
e
t
GT
P.
Ce
d
o
m
ai
n
e est
l
arge
m
e
nt
di
s
tribu
é a
u
se
i
n
d
es gè
n
es
R
cl
o
n
és et
es
t
co
mp
osé no
t
a
m
ment
d
e
d
iffére
nt
s
m
o
ti
fs
d
e ry
p
e
kin
ase. L
'
analog
i
e
d
e ce
d
o
m
a
in
e avec des pro
t
é
in
es a
nim
a
l
es
p
ote
nt
ie
ll
ement i
m
p
li
q
u
ées da
n
s
l
es p
h
é-n
o
m
è
n
es
d
e
m
o
re c
e
llul
a
ir
e a
p
op
totiqu
e
a con
duit
à
l
a
n
ot
i
o
n d
e
d
o
m
ai
n
e N
B-ARC [
1
4]
.
Deux autres
d
o
m
ai
n
es
,
LZ et
T
IR
,
m
o
in
s
fr
éq
u
e
nt
s a
u
se
in d
es gè
n
es
R
i
d
e
nti
fiés et gé
n
é
r
ale
m
e
nt
assoc
i
és aux
p
rod
ui
ts N
B
S
-LRR
,
o
nt
été
mi
s e
n
év
i-d
e
n
ce.
L
e
d
o
m
ai
n
e LZ
(Leucine Zipper)
N Apoplasme Membrane plasmique
..
~ Ser/Thr kinase cÎxa21 Cytoplasme Figure 1. Représen -tation schématique et localisation pos -sible des domaines protéiques conser -vés au sein des pro -duits de gènesR
clonés [12, 16]. Figure 1. Schematic representation and putative location of conserved protein domains of cloned R genes.es
t
co
nnu p
o
u
r avo
ir un r
ô
l
e
d
a
n
s
l'h
o
m
o o
u l
'
h
é
c
éro
dim
é
ri
sa
ci
o
n d
es
p
ro
-t
é
in
es (
1
5
], c
a
ndi
s
qu
e
l
e
d
o
main
e T
IR
(To
lf l
n
t
er
l
eukin
R
ece
p
tor)
p
rése
n
ce
d
'
imp
o
rt
a
nc
es
h
o
molo
g
i
es
d
e sé
qu
e
n
ces
a
v
e
c l
es
dom
a
in
es
d
e ré
cep
te
ur
s
pro-t
é
iqu
es
i
s
ol
és
d
e
l
a
dro
s
ophil
e (
r
éce
p
te
ur
T
o
ll
) e
t d
e
l
'
homm
e
(r
éce
p
te
ur in
ce
rl
e
u-kin
e
-1
)
(1
2
]
.
Sur
l
a
b
a
se d
e ces
analo
g
i
es,
un r
ô
l
e
dan
s
la
casca
d
e
d
e s
i
g
nali
sat
i
o
n
ce
llul
a
ir
e es
t
gé
n
é
ral
e
m
e
nt
a
ttribu
é a
u
d
o
m
a
in
e T
IR.
Classification des gènes
R
Qu
a
rr
e
cl
asses
d
e
g
è
n
es
R
o
ne
é
t
é
d
é
fini
es
s
ur
l
a
b
ase
d
es
diff
é
r
e
nt
es assoc
i
a
ti
o
n
s
d
es
dom
a
in
es
cons
er
v
és (
LRR
, N
B
S,
L
Z,
TIR
e
t S
e
r/Thr kina
se)
au
se
in d
es
p
ro-duit
s
d
e gè
n
es
R
(ta
bl
ea
u
2)
(1
2,
16].
L
a
pr
e
mi
è
r
e
cla
sse
d
e gè
n
es
R
co
d
e
r
a
it
p
o
ur d
es
prot
é
in
es cy
topl
as
miqu
es
d
e
ty
p
e réce
pt
e
u
r. Ces gè
n
es
pr
ése
nt
e
nt
à
l
a
fo
i
s
un dom
a
in
e
LRR
à
l
'ex
tr
é
mit
é 3
'
e
t
un d
o
m
ai
n
e
NB
S à
l
'ex
tr
é
mit
é 5
'
. Ces
gè
n
es
NBS-LRR
c
on
s
cicu
e
nc
l
a
prin
c
ipal
e
cl
asse
d
e gè
n
es
R
clon
é
s ju
s
qu
'à
pr
ése
nt.
A
u
se
in d
e ce
tt
e
cl
asse,
d
es
produit
s
d
e
ce
rt
a
in
s gè
n
es
pr
ése
nt
e
nt
e
n po
s
iti
o
n
amin
o-ce
rmin
a
l
e
l
es
moti
fs T
IR
o
u
LZ,
e
n plu
s
d
es
dom
a
in
es
LRR
e
t NB
S.
La d
e
uxi
è
m
e
classe regroup
e
l
es gè
n
es
Cf2
(1
7
)
e
t
Cf9
(18) d
e
la tom
a
t
e e
t l
e
gè
n
e
H
S
J
Pro
-l
d
e
l
a
b
e
tt
e
r
ave s
u
c
ri
è
r
e
(
19)
.
L
es
p
ro
duit
s c
od
és
p
a
r
ces gè
n
es
pr
ése
n
te
nt un d
o
maine LRR
ex
t
race
llu
-l
ai
r
e,
un dom
ai
n
e
tran
s-
m
e
mbran
ai
r
e e
c
un
e co
urre
r
ég
ion c
yt
opl
as
miqu
e.
Tableau 2
Les tro1S1eme et
qu
atriè
m
e classes
n
e
co
nti
e
nn
e
nt
po
u
r l'i
n
stan
t
q
u
'
un
se
ul
é
l
é
m
e
nt. L
a
t
ro
i
s
i
è
m
e
cl
asse es
t r
e
pr
é
-se
nt
ée
p
ar
l
e gè
n
e
Pto
(20). Ce gè
n
e
co
d
e
un
e
p
ro
t
é
in
e
kin
ase
d
e ty
p
e sér
i-n
e/
thr
é
onin
e
.
Bi
e
n qu'
a
u
c
un
e é
cude
n
'
ait p
e
rmi
s
d
e
m
o
ntr
e
r un
e assoc
i
a
ti
o
n
dir
ecte e
ntre l
es
d
eux
prot
é
in
es,
l
es
pro-duit
s
d
es
g
è
n
es
P
to
e
t
P
rf (
d
e
cl
asse
N
B
S-
LRR
) so
nt
to
u
s
l
es
d
e
u
x ex
i
gés
p
o
ur qu
'
il
y ai
t
ex
p
ressio
n d
e
l
a
r
és
i
s
-t
a
n
ce a
pr
ès
inf
ec
ti
o
n p
a
r
P.
syringae
ex
pr
iman
t l
e gè
n
e AvrPto
[2
1
)
.
Le gè
n
e
Xa2
1
c
on
s
titu
e
l
a
qu
a
tri
è
m
e
cl
asse
(
22)
.
Un dom
a
in
e
LRR
ex
t
race
llul
a
ir
e,
un
e
rég
ion tr
ans
-m
e
mbr
a
n
a
i
re e
t un d
o
m
ai
-n
e
kin
ase sé
rin
e/
thr
éo
nin
e cy
t
o
pl
as
-miqu
e
ont
é
t
é
id
e
n
t
ifi
és
au
se
in d
e
l
a
prot
é
in
e Xa2
l.
L
a s
tru
c
tur
e
d
e ce
tt
e
prot
é
in
e s
u
ggè
r
e,
d
'
un p
o
int d
e v
ue
év
olutif
,
un
e
rel
at
i
o
n
e
ncr
e
l
es
prot
é
in
es
d
e ty
p
e
LRR
(Cf) e
c
ce
ll
es
d
e ty
p
e
kin
a
-se (
Pro
)
.
E
nfin
,
il
co
n
vie
nt d
e
m
e
nti
o
nn
e
r qu
e
certa
in
s gè
n
es
d
e rés
i
sta
n
ce
n
'ap
p
art
i
en-n
e
n
t à a
u
c
un
e
d
e ces
qu
a
rr
e
cl
asses.
C'es
t l
e cas,
p
ar exe
mpl
e
du
gè
n
e
H
m
l
,
c
h
ez
l
e
m
a
ï
s,
qui
co
n
t
r
ô
l
e à
l
a
foi
s
l
a
r
és
i
s
tan
ce a
u
c
h
a
mpi
g
non patho
gè
n
e
Coc
hl
iobo
l
us carbonum
r
ace
1
e
t
l
'ex
pr
ess
ion d
'
un
e
t
ox
in
e
r
é
du
c
t
as
e.
Ce
rc
e
r
é
du
c
t
ase
pr
ése
nc
e
l
a
c
a
p
ac
it
é à
in
ac
ti
ve
r un
e tox
in
e
du
c
h
a
mpi
g
n
o
n
p
a
tho
gè
ne
.
H
m
l
se
diff
é
r
e
n
c
i
e
de
s
gè
n
es
R pr
écé
d
e
mm
e
nt
c
ir
és
p
a
r
l
e fa
ir
qu'
a
ucun
com
p
osé
Avr
n'
es
t impliqu
é
d
a
n
s
l
a
d
égra
d
a
ti
o
n d
e
l
a tox
in
e
p
a
r
l'
ac
ti
v
it
é
r
é
du
ctase (23).
Origine de la spécificité
des gènes
R
L
'
é
tud
e des
b
ases
m
o
l
éc
ul
aires
d
e
l
a
spé-c
i
fic
i
té de
r
eco
nn
a
i
ssa
n
ce
r
este e
n
core
un
o
bj
ec
tif m
a
j
e
u
r
d
es
r
ec
h
er
ch
es
d
e
l'in
te
r
-act
i
o
n
gè
n
e-à
-
gè
n
e
(1
3).
L
e
d
o
m
a
in
e
L
RR
es
t fr
é
qu
e
mm
e
nt impliqu
é
dan
s
l
es
int
e
r
ac
ti
o
n
s
p
ro
t
é
in
e
-prot
é
in
e [24
)
co
mm
e,
p
a
r
exe
mpl
e,
l
ors
d
'
int
e
r
ac
ti
o
n
s
spéc
ifiqu
es e
ntr
e
d
es
r
éce
p
te
ur
s
d
'
h
o
r-m
o
n
es e
t l
e
u
r
li
ga
nd
s g
l
ycopro
t
é
i
q
u
es
c
h
ez
l
es
m
amm
i
fères (25)
. Il
se
mbl
e
r
a
it
q
u
e
l
a
r
ég
i
o
n LRR d
es
p
ro
duit
s
d
e gè
n
es
R
soi
t
force
m
e
n
t
li
ée à
l
a s
p
écific
it
é
d
e
l'
inc
e
r
acc
i
o
n
gè
n
e-à-gè
n
e (2
6).
L
'a
n
a
l
yse
d
es
t
a
u
x
d
e s
ub
s
tiruti
o
n
s
nucl
éo
cidiqu
es a
u
se
in d
es sé
qu
e
n
ces
LRR d
es gè
n
es
Cf4 e
t
Cf 9
(gè
n
es
d
e
t
y
p
e
LRR
) s
u
ggè
r
e
qu
e ces
m
o
ti
fs
s
tru
c
tu
raux so
nt
s
oumi
s à
un
e
p
ress
i
o
n
d
e d
i
ve
r
s
ifi
cat
i
o
n
[27], ce
qui p
araî
t
co
h
ére
nt p
o
u
r
un
e
r
é
g
i
on
ass
ur
a
nt
l
a
d
é
t
er
min
a
ti
on
d
e
l
a s
p
éc
i
fic
i
ré
. D
es
o
b
se
r
vat
i
o
n
s s
imil
a
i
res o
ne
é
t
é e
ff
ec-tu
ées
d
a
n
s
l
e cas
d
e
la
fa
mill
e
du
gè
n
e
Xa2
1
(28]. Pa
r
a
ill
e
ur
s,
un
e
imp
o
rt
a
n
te
é
tud
e, co
mbin
a
n
t co
mp
a
r
a
i
so
n d
es
sé
qu
e
n
ces
d
e
di
ffé
r
e
nts
a
ll
è
l
es e
t an
a
l
yse
fo
n
c
ti
o
nnell
e
d
'al
l
è
l
es c
him
é
riqu
es
c
h
ez
d
es
pl
a
nt
es
tr
a
n
sgé
niqu
es, a é
t
é
m
e
n
ée
a
u locu
s
L
du
lin
[2
9
]. Ce
locu
s es
t
co
mpo
sé
d
e
1
3
all
è
l
es e
t
co
d
e
d
es p
ro
-duit
s
d
e
t
y
p
e
LRR
-N
B
S
-
T
IR qui
o
nt
é
t
é sé
qu
e
n
cés e
t
a
n
a
l
ysé
s. Il
a
pp
araî
t
q
u
e
d
e
u
x
d
es pro
duit
s
d
é
duit
s,
L
6 et
L
l l
,
se
di
ffé
r
e
n
c
i
e
nt s
e
ul
e
m
e
nt
a
u
ni
vea
u d
e
l
e
ur r
ég
i
o
n
L
RR t
a
ndi
s
q
ue
Les quatre classes de gènes
R
définies sur la base des domaines protéiques conservés
Classe Gène de résistance Plante Agent pathogène Gène d'avirulence Structure Références
RPS2 Arabidopsis Pseudomonas syringae pv. tomato AvrRpt2 LRR-NBS-LZ [71, 72]
RPM1 Arabidopsis P. syringae pv. macu/icola AvrRpm1, avrB LRR-NBS-LZ [73]
Prf Tomate P. syringae pv. tomato AvrPto LRR-NBS-LZ [74]
Mi Tomate Meloidogyne spp.
?
LRR-NBS-LZ [75]N
Tabac Tobacco mosaic virus TMV LRR-NBS-TIR [76]L6 Lin Melampsora lini AL6 LRR-NBS-TIR [77]
M Lin Melampsora lini AM LRR-NBS-TIR [78]
RPP5 Arabidopsis Peronospora parasitica AvrPp5 LRR-NBS-TIR [79]
l2C Tomate Fusarium oxysporum
?
LRR-NBS [80]Rp1-D Maïs Puccinia sorghi
?
LRR-NBS [81]Il
Cf-9 Tomate Cladosporium fulvum Avr9 LRR [18]Cf-2 Tomate
C.
fulvum Avr2 LRR [17]HSJPf0-1 Betterave Heterodera schachtii
?
LRR [19]Ill
Pto Tomate P. syringae pv. tomato AvrPto Protéine Kinase [20]IV Xa21 Riz Xanthomonas oryzae pv. oryzae
?
LRR-PK [22]les deux produits déduits L6
et
L7 ne
diffèrent que par
l
eur
région TIR.
Ainsi, la région TIR pourrait,
comme
l
a
région LRR
,
intervenir dans la
spéci-ficité de
cette
interaction.
Dans
l
e cas
de
l
a
kinase Pto qui ne
pré-sente
ni domaine LRR ni domaine
TIR
,
l
a suppression
d
'
un
e
thréonine
en
position 204
se
traduirait par la perce
de la
spécific
it
é
de l'interaction
Avr
Pto!Pto [30].
Gènes R: des locus
complexes
De
s
études génétiques et
moléculaires
ont révélé
une
organisation génomique
particulière des
gènes
de résistance. La
plupart d
'e
ntre
eux sont génétiquement
étroitement associés à
d
'
autres
gènes o
u
séquences
homologues
et conscicuent
des
locu
s comp
l
exes
ou
clusters [31, 32].
Dans la bibliographie
concernant
l
es
gènes
R
,
le terme d
'a
llèle désigne
l
es
formes d
'
un
gène à
un lo
cus
donné mais
aussi,
dans de nombreux
cas,
les
formes
présentes
aux gènes apparentés
adja-cents.
De
l
a
même manière,
à
un locus
R peuvent
correspondre
plusieurs
gènes.
Classiquement,
l
e
qualificatif d
'
homo-lo
gues est attribué aux gènes
présentant
des
séquences et,
par déduction
,
des
fonctions
apparentées.
Deux types de
gènes
homologues
sont communément
distingués. Les gènes qui
,
au sein
d
'es
pèces différentes,
assurent
l
a
même
fonction
et
occupent une position
cor-respondante
sur
l
es chromosomes sont
appe
l
és gè
n
es ort
hologu
es. Au se
in
d
'
une
même
espèce,
l
es gènes apparentés
non-a
ll
è
l
es
résultant de
dupli
cations sont
dits
paralogues.
Locus
à
multiples
spécificités de résistance
La majorité des
l
ocus aujourd'hui
réper-toriés présentent de multiples
spécifici-tés.
Au
niveau de
ces
l
ocus, on
retrouve
de nombreux allèles qui
confèrent
l
a
résistance à
une race
spécifique
de l
'ag
ent
pathogène
co
n
sidéré
(figure 2).
Par
exem
pl
e, c
h
ez
l
e
lin,
l
a
résistance
à
l
a
rouille
Melampsora
fini
est assurée
par
au
moins
30 a
ll
è
l
es
r
egroupés en 7
l
oc
u
s
pouvant présenter des
organisat
i
ons
variées.
Par
exemp
l
e,
le
l
ocus
L
corres-pondrait
à
un
gène
unique présentant
au
moins 13 allèles
(L,
LI
à
LI
1
et
LH)
correspondan
t
à
13
spéc
ifi
c
it
és
[29],
tan-di
s
que
le
l
ocus
M
serait composé
d
'e
nvi-ron 15
gènes e
n
c!usters et assurant 7
spé-cificités connues [33].
Co-localisation de gènes
de résistance
à
différents
agents pathogènes
Des
lo
cus
de résistance, assurant une
résistance
spécifique
dirigée contre
diffé-rents agents pathogènes, peuvent
être
co-l
ocal
isés
sur
un même fragment
chromo-som
iqu
e.
De tels
lo
cus
ont
été
répertoriés
au sein
de diverses
espèces
végétales telles
que
l
a
laitue
[34],
le blé
[35] ou encore
la tomate. Par
exemple
chez
la tomate
,
il
a été
montré que
,
sur
une région d
'e
nviron
20 cM
à
l
'ex
trémi-té
du bras
court
du
chromosome
6,
sont
regroupés : deux
spécificités
de résistance
à
Cladosporium falvum (Cf2 ec Cf5)
[36]
;
l
e gène
Mi, gène
de résistance
au
nématode
Meloidogyne
incognita
[37]
;
l
e
gène
de
résistance
Ol-1
à
Oidium
IJco-pers
icum
[38) ;
le
gène
Meu] responsable
de
l
a
résistance
au
puceron
Macrosiphum
euphorbiae [39] et
l
e gène
Ty-1
de
to
l
é-rance au TYLCV
(Tomato Yellow Leaf
Curl Virus) [40].
Multiples copies
de séquences analogues
organisées en
clusters
Récemment
,
un
grand
nombre de
séquences
présentant des
sim
il
ar
it
és
avec
les
gènes
R
clonés
ont été
mises
en évidence au sein
des
génomes
de
plantes
: on
parle d'analogues de
gè
n
es
de résistance
(ou
RGA,
Resistance
Gene
Analogs).
Ces
RGA one
été
l
e
plus
souvent
obtenus
et caractérisés
s
uit
e à
des
amp
lifi
cations
in vitro
(PCR)
utilisant des
amorces
oligonu-cléotidiques dégénérées
correspondant
à
des domaines
conservés (c'est-à-dire
NBS) des
gènes
de
résistance.
La
plu-part de
ces
RGA
sont
organisés en
clusters ec co-lo
calisés avec
des
gènes
de résistance
connus
[35,
4
1-
49]. Une
celle organisation
correspond
à
un
e
nsemble de
séquences
répétées en
tandem. Les gènes de résistance de
types
NBS-LRR
ou LRRJkinase et
les
séquences
qui
l
eur sont apparentées
constituent ainsi
des
fami
ll
es
multigé-niques majeures
au sein
des
génomes
de plantes [50).
Cahiers Agricultures 2000
;
9
:
301-9
A. 1 M m1_. ---c::J---m--
- -
-If- -CJ-- -2 m M1-' --D---D---D-
-
-1-f- -
- -
-B. 1. 2. 3.15 gènes Men tandem gène L unique
L2
-
L6
~~~-'[:=J~~~~-Figure 2. Représentation schématique des
structures génétiques des locus L et M de
résistance à la rouille chez le lin. Les
rec-tangles correspondent aux régions codantes apparentées (33).
A. Le locus M: 1. la spécificité M est présen-te au sein de lignée du lin Forge; 2. la
spéci-ficité M1 au sein de lignée du lin Williston
Brown. Les positions relatives de M et M1
sont arbitraires.
B. Le locus
L
:
les 3 allèles, ici présentés L2,L6 et L 10, ont été respectivement identifiés
au sein des lignées de lin Stewart, Birio et
Bolley Golden.
Figure 2. Schematic representation of genetic structures at the L and M rust resistance loci in
flax. Rectangles indicate related coding
sequences.
A. The M locus complex : 1. M specificity pre-sent in the Forge flax line; 2. M 1 specificity in
the Williston Brown flax line. The relative posi-tions of M and M1 are arbitrary.
B. The L locus: alleles L2, L6 and L 10 are
pre-sent respectively in the Stewart, Birio and Bol-ley Golden flax lines.