THÈSE
Pour l'obtention du grade de
DOCTEUR DE L'UNIVERSITÉ DE POITIERS UFR des sciences fondamentales et appliquées
Institut de paléoprimatologie, paléontologie humaine : évolution et paléoenvironnements - IPHEP (Poitiers)
(Diplôme National - Arrêté du 7 août 2006)
École doctorale : Sciences pour l'environnement - Gay Lussac (La Rochelle) Secteur de recherche : Terre solide et enveloppes superficielles
Présentée par :
Salamet Mahboubi
Les rongeurs du miocène supérieur et terminal d'Afrique
nord-occidentale : biochronologie, magnétostratigraphie,
biogéographie et paléoenvironnements
Directeur(s) de Thèse :Jean-Jacques Jaeger, Mouloud Benammi Soutenue le 12 décembre 2014 devant le jury
Jury :
Président Patrick Vignaud Professeur des Universités, Université de Poitiers
Rapporteur Sevket Sen Directeur de recherche, MNHN, Paris
Rapporteur Lawrence J. Flynn Assistant director, Peabody Museum, Harvard University Membre Jean-Jacques Jaeger Professeur émérite, Université de Poitiers
Membre Mouloud Benammi Ingénieur de recherche CNRS, Université de Poitiers Membre Jacques Michaux Directeur d'étude honoraire, Université de Montpellier Membre Pablo Peláez-Campomanes Investigador, Museo nacional ciencias naturales, Madrid
Pour citer cette thèse :
Salamet Mahboubi. Les rongeurs du miocène supérieur et terminal d'Afrique nord-occidentale : biochronologie, magnétostratigraphie, biogéographie et paléoenvironnements [En ligne]. Thèse Terre solide et enveloppes superficielles. Poitiers : Université de Poitiers, 2014. Disponible sur l'Intranet de l'Université de Poitiers <http://theses.univ-poitiers.fr>
THESE
Pour l’obtention du Grade de DOCTEUR DE L’UNIVERSITÉ DE POITIERS (Diplôme National – Arrêté du 7 août 2006)
Ecole doctorale : Sciences pour l’Environnement – Gay Lussac Secteur de Recherche : Terre Solide et Enveloppes Superficielles
Discipline : Paléomagnétisme et Paléontologie des Vertébrés
Présentée par :
Salamet MAHBOUBI
LES RONGEURS DU MIOCÈNE SUPÉRIEUR ET TERMINAL D’AFRIQUE
NORD-OCCIDENTALE : BIOCHRONOLOGIE, MAGNÉTOSTRATIGRAPHIE,
BIOGÉOGRAPHIE ET PALÉOENVIRONNEMENTS
Sous la direction de :
Jean-Jacques JAEGER, Professeur Émérite Université de Poitiers Mouloud BENAMMI, Ingénieur de Recherche CNRS
Soutenue le 12 décembre 2014
Jury
Lawrence J. FLYNN Directeur adjoint, Université de Harvard Rapporteur Sevket SEN Directeur de Recherche, MNHN, Paris Rapporteur Jacques MICHAUX Directeur d’Étude, EPHE, Montpellier Examinateur Pablo PELÁEZ-CAMPOMANES Directeur de Recherche, MNCN, Madrid Examinateur Patrick VIGNAUD Professeur, Université de Poitiers Examinateur Jean-Jacques JAEGER Professeur Émérite, Université de Poitiers Directeur de thèse Mouloud BENAMMI Ingénieur de Recherche CNRS, Poitiers Directeur de thèse
«En vérité, le chemin importe peu, la volonté d'arriver suffit à tout.»
Remerciements
La Thèse est la dernière étape conduisant au monde de la recherche. C’est un travail enrichissant qui nécessite un investissement personnel intense. C’est cependant la partie que je trouve la plus agréable à écrire. Je tiens à remercier ici toutes les personnes qui m’ont soutenue, qui m’ont encouragée, et qui ont contribué de près ou de loin à l’aboutissement de ce travail.
Je tiens tout d'abord à exprimer ma sincère gratitude à mon directeur de thèse Jean-Jacques JAEGER. Je lui dois beaucoup pour l’initiation dans le domaine de la paléontologie, pour le temps et les efforts consacrés durant la conduite de ce travail, et pour tout ce qu'il m'a appris ; merci également pour votre confiance et votre grande générosité qui a toujours montré au-delà du domaine professionnel. Grâce à vous j’ai enfin réalisé mon rêve d’enfance.
Je tiens à exprimer mes sincères remerciements à Mouloud BENAMMI, qui a également dirigé ce travail. Il m’a initié aux méthodes et techniques du paléomagnétisme tout en assurant mon encadrement sur le terrain. Sa confiance m’a permis d’avancer. Qu’il soit assuré de ma profonde gratitude. Les résultats présentés ici sont les conséquences de son aide précieuse et son soutien permanent durant la préparation de cette thèse.
J’adresse mes remerciements à Sevket SEN et Lawrence J. FLYNN, d’avoir non seulement accepté de faire partie de mon jury, mais aussi d’avoir rapporté mon travail. Je tiens également à remercier Jacques MICHAUX, et Pablo PELÁEZ-CAMPOMANES et Patrick VIGNAUD pour m’avoir fait l’honneur de juger ce travail.
Je tiens à remercier très respectueusement Michel BRUNET pour ses encouragements et de ses visites permanentes dans mon bureau même durant les week ends. Je lui exprime mes sentiments les plus vifs et les plus sincères.
Ma sincère reconnaissance va tout spécialement à Patrick VIGHAUD, Directeur de l’iPHEP, pour sa disponibilité et pour ses nombreuses interventions pour aplanir toutes les difficultés en me suggérant des solutions. Sa présence dans le jury est un honneur pour moi.
Je remercie également Sylvie PÉREZ de l’administration, Sabrina BIAIS et Boniface KOKOH de l’Ecole doctorale Gay Lussac. Mes remerciements s’adressent également à Anne BRUNELLIÈRE, pour son efficacité dans la recherche d’articles et sa grande gentillesse. Merci à Emile BERE pour la microscopie électronique. Grand merci à François Baty-Sorel et François Lecontte.
Tous mes remerciements vont à l’ensemble du personnel de l’iPHEP pour leur accueil depuis le Master 2. Par leur bonne humeur et pour leurs encouragements cruciaux pendant toute la durée de cette thèse, et particulièrement ces dernières semaines. Cette ambiance de travail m’a permis de supporter les moments de démotivation et de découragement: Ghislaine, Carine, Cécile, Olga, Géraldine, Stéphane, Xavier, Jean-Renaud, Olivier, Franck, Sabine, Vincent, Guylaine, Guillaume, Yaowalak…. Merci!. Une mention spéciale à Renaud : merci pour ton soutien moral durant les dernières semaines. Merci à Gildas Merceron pour ses discussions constructives.
Je remercie très vivement les chercheurs que j’ai rencontrés pendant mon séjour en France, parmi eux je cite : Rodolphe TABUCE, Laurent MARIVAUX, Fabrice LIHOREAU, Monique VIANEY-LIAUD et Pauline COSTER.
réservé un accueil chaleureux et une aide indispensable. Je cite la famille BENAMMI et la famille TERMIDI au Maroc : merci pour votre accueil chaleureux.
J’exprime une pensée respectueuse toute particulière aux « Hommes du désert » qui m’ont accompagné lors des missions de terrain effectuées à Oued Mya (Sahara Central) : Cheikh MAMMERI, Mustapha MAMMERI Abdelhamid GABANI, Ali BOUGUERRA, Bahous SLAMET, Mohamed BELKHEIRI et Okacha KHEDOUM. Merci pour votre patience et pour le verre de thé du soir qui m’a permis de supporter les nuits longues et froides du Tademaït. Sans votre assistance et votre aide, ce site paléontologique d’Oued Mya serait perdu à tout jamais. Par la même occasion je remercie Taïb BOUIMA, Maître de Conférences à l’Université de Sétif, qui m’a réservé un accueil familial tout en me faisant profiter de son expérience sur le terrain pour résoudre un certains nombre de problèmes liés à la géologie du bassin d’El Eulma.
Je voudrais aussi exprimer mes remerciements aux chercheurs et personnel du Laboratoire de Paléontologie de l’Université d’Oran de m’avoir accueilli pendant 3 mois lors de ma première année de thèse et en me fournissant tout le matériel nécessaire pour les manipulations.
Je n’oublierai jamais ceux qui m’ont entouré par leur soutien, leur compassion et leurs encouragements, ami(e)s et collègues, en particulier Anne, Florian, Dimitri, Dom, Alice, Bouziane, Meriem, Soria, Ryma, Amira et Sohee.
Un grand merci à Antoine SOURON et Ghislain THIERY pour leur aide et leur disponibilité permanante et notamment pour la lecture des différentes parties de cette thèse. Qu’ils trouvent ici ma profonde reconnaissance.
Merci infiniment à Tiphaine COILLOT pour ton précieux soutient sous toutes ses formes durant cette dure période, merci pour ta présence à mes côtés et pour m’avoir apporté tous les soins dont j’avais besoin durant cette péride de stress permanent.
Je suis très reconnaissante et redevable envers EURL-TASSINA (El Bayadh, Algérie) et l’iPHEP qui ont soutenu financièrement les travaux de cette thèse.
Je ne peux terminer sans remercier pleinement mes parents, mes frères Yasser, Abdou et ma sœur Asma pour leur patience et leur aide inestimable. Qu’ils trouvent dans ce travail le gage de mon amour et de ma reconnaissance. Je dois beaucoup à mon père qui m’a fait découvrir ce beau métier de géologue dès mon plus jeune âge lors des missions de terrain que j’ai accomplies en sa présence sur l’Atlas saharien, la Saoura, le Tademaït et le Tanezrouft.
Table des matières
Introduction ... 3
Chapitre I. Matériels et Méthodes ... 7
1 Introduction ... 9
2 Localités et origines du matériel étudié ... 9
3 Prélèvement du matériel ... 10
3.1 Mesures ... 11
3.2 Taxonomie paléontologique ... 11
3.3 Nomenclature ... 11
3.4 Photographie ... 15
4 Biochronologie du Miocène continental en Afrique du Nord ... 15
5 Magnétostratigraphie ... 20
5.1 Prélèvement et préparation des échantillons ... 21
5.1.1 Échantillonnage ... 21
6 Les différents modes d’acquisition de l’aimantation dans les roches sédimentaires ... 22
6.1 Acquisition de l’aimantation rémanente (ARN) ... 22
6.1.1 L’Aimantation Thermorémanente (ATR) ... 23
6.1.2 L’Aimantation Rémanente Isotherme (ARI) ... 23
6.1.3 L’Aimantation Rémanente Chimique (ARC) ... 23
6.1.4 L’Aimantation Rémanente Visqueuse (ARV) ... 23
6.1.5 L’Aimantation Rémanente Détritique (ARD) ... 23
6.2 Minéralogie magnétique ... 24
6.3 Méthodes d’identification des minéraux ferromagnétiques (Méthode de Lowrie, 1990) 25 6.3.1 L’acquisition de l’aimantation rémanente isotherme (ARI) ... 25
6.3.2 Mesure de l’aimantation rémanente naturelle (ARN) ... 25
7 Traitement des données ... 27
7.1 Acquisition et visualisation des données ... 27
7.2 Tests paléomagnétiques ... 28
Partie 1 : Etude magnétostratigraphique et paléontologique des sites continentaux néogènes d’Algérie ... 31
2 Cadre géographique et géologique ... 35 3 Description de la coupe ... 38 3.1 Formation I ... 38 3.2 Formation II ... 39 4 Données biostratigraphiques ... 41 5 Magnétostratigraphie ... 43 5.1 Prélèvements paléomagnétiques ... 43
5.2 Traitement magnétique et identification des minéraux ... 43
5.3 Acquisition de l’aimantation rémanente isotherme (ARI) ... 44
5.4 Désaimantation thermique de l’ARI ... 45
6 Analyse de l’aimantation rémanente naturelle (ARN) ... 47
6.1 Mesure de l’aimantation rémanente naturelle ... 47
6.2 Analyse de l’aimantation rémanente naturelle ... 47
7 Discussion ... 54
7.1 Succession des polarités magnétiques ... 55
7.2 Corrélation avec l’échelle des polarités géomagnétiques ... 57
7.3 Taux de sédimentation ... 59
7.4 Implications structurales... 60
8 Conclusions ... 61
Chapitre III. Nouvelles données chronologiques relatives au bassin de la Tafna : Combinaison des données bichronologiques et magnétostratigraphiques. ... 63
1 Introduction ... 66
2 Geological setting ... 67
2.1 Regional description of the basin infill ... 67
2.2 The Djebel Guetaf section ... 67
3 Systematic paleontology ... 70
4 Paleomagnetic analyses ... 76
4.1 Sampling and laboratory procedures ... 76
4.2 Isothermal Remanent Magnetization (IRM) ... 76
4.3 Natural Remanent Magnetization (NRM) analysis ... 78
5 Correlations and discussion ... 83
6 Conclusions ... 86
Partie 2. La faune de micromammifères du gisement d’Afoud (Bassin d’Aït Kandoula, Maroc) : Nouvelle preuve des échanges fauniques afro-ibériques pendant le Messinien. . 89
Chapitre IV. Systématique ... 91
1 Introduction ... 93
2 Cadre géographique et géologique du Bassin d’Aït Kandoula ... 93
3 Description de la coupe ... 96
4 Étude systématique ... 98
4.1 Description du matériel du gisement d’Afoud « AF12-1 » ... 99
4.2 Description du matériel du gisement d’Afoud « AF12-2 » ... 105
4.3 Description du matériel du gisement d’Afoud « AF12–3 » ... 152
5 Conclusions sur l’étude paléontologique des trois nouveaux gisements fossilifères ... 154
Chapitre V. Biochronologie ... 155
1 Introduction ... 157
2 Position stratigraphique des gisements de rongeurs d’Afoud... 157
3 Une approche d'âges absolus des gisements étudiés ... 160
Chapitre VI. Essai de reconstitution paléoenvironnementale ... 163
1 Introduction ... 165
2 Essai de reconstitution paléoenvironnementale à partir des assemblages de rongeurs ... 166
2.1 Comment les assemblages de rongeurs permettent-ils de caractériser les paléoenvironnements? ... 166
2.2 Remarques sur la taphonomie les sites échantillonnés ... 167
2.3 Composition quantitative des associations de micromammifères ... 168
2.4 Caractérisation bioclimatique des espèces fossiles ... 170
2.5 Quel signal paléoenvironnemental les assemblages de rongeurs ont-ils enregistré ? 172 2.6 Estimation des paléotempératures ... 175
3 Implications palynologiques et géochimiques ... 175
3.1 Les données polliniques ... 175
3.2 Les données géochimiques ... 176
4 Comparaisons avec d'autres sites continentaux mio-pliocènes à rongeurs ... 178
Chapitre VII. Paléobiogéographie ... 183
1 Introduction ... 185
2 Facteurs de migration ... 186
3 Migrations des mammifères et des micromammifères pendant le Miocène supérieur... 186
3.1 Migrations dans le sens Est–Ouest ... 186
3.2 D'autres migrations de micromammifères durant le Turolien supérieur ... 188
4 Les échanges fauniques entre le Maghreb et l’Espagne et le problème des migrations ... 190
4.2 Échanges fauniques entre l’Espagne et le Maghreb ... 195
5 Fin de la voie maritime en Méditerranée nord-occidentale: un temps pour un passage terrestre éphémère ... 196
5.1 Première migration afro-ibérique dans le sens Sud–Nord à 6.3 Ma... 199
5.2 Deuxième migration entre 6.12 Ma et 6.2 Ma ... 201
5.3 Troisième migration entre 5.9 et 5.3 Ma ... 201
6 Conclusions ... 202
Conclusions et perspectives ... 203
Bibliographie ... 209
Introduction
Le Miocène supérieur est une période de grands changements climatiques, environnementaux et fauniques. Les registres fossiles, dans de nombreuses régions, témoignent que le climat pendant la fin du Miocène (de 11.61 Ma à 5.33 Ma) a été beaucoup plus chaud et/ou humide qu'aujourd'hui (Hilgen et al, 1995 ; Pound et al., 2011, 2012). Parallèlement à ces changements climatiques on assiste, sur les continents, à l’émergence des plantes en C4 (Quade et al., 1989 ; Cerling et al., 1993 ; Quade et Cerling, 1995), qui sont enregistrées dans les basses latitudes vers 7 Ma et 8 Ma, puis dans les latitudes moyennes vers 5 Ma (Cerling et al., 1997). Par exemple, les forêts tempérées chaudes ont prospéré dans des régions circum-boréales (Denk et al, 2005). Les prairies existaient dans des régions actuellement désertiques (par exemple, la péninsule arabique, Kingston et Hill, 1999 ; le désert du Sahara, Vignaud et al., 2002). L’apparition de ces graminées en C4 est fréquemment interprétée comme étant liée à l’ouverture des milieux. En outre, une calotte glacière à l’Antarctique a existé tout au long de la fin du Miocène (Lewis et al., 2008).
La fin du Miocène est aussi une période où des réorganisations tectoniques importantes ont eu lieu : par exemple, le soulèvement tectonique des Himalayas (Harrison et al., 1992), celui des Andes (Garzione et al, 2008), celui des Plateaux d’Afrique orientale, celui des Alpes, et la fermeture du détroit de Gibraltar pendant la crise de salinité messinienne (Hsü et al., 1973). Cette période a aussi connu une transition entre les faunes de mammifères archaïques du Miocène moyen et les faunes d’aspect moderne (Hill, 1995 ; Le Fur et al., 2009). L’arrivée des Hipparioninés autour de 11 Ma, avec une grande phase de dispersion autour de 10.7 Ma. vers l’Eurasie et l'Afrique (Sen, 1986 ; Garcès et al., 2003) a marqué un événement majeur pendant cette période. À partir de 9.5 Ma les espèces de rongeurs semblent indiquer des environnements plus instables avec une baisse de la longévité des espèces (Flynn et al., 1995). L’aridité et la saisonnalité ont aussi augmenté durant cette période, cette augmentation est marquée dans les milieux tropicaux par l’expansion des plantes en C4 (Pound et al., 2011, 2012).
C’est dans ce cadre de bouleversements, d’instabilité climatique, et de renouvellement faunique que les hominidés ont émergé. La découverte du plus ancien hominidé au Tchad, Sahelanthropus tchadensis (Brunet et al., 2002) à 2500 km à l’ouest du Rift, vient clôturer définitivement la théorie de l’ « East Side Story » (Coppens, 1983). Cette découverte a engendré de nouvelles problématiques concernant les paléoenvironnements africains et les connexions fauniques entre les différentes zones géographiques au sein de ce continent. Un réseau hydrographique important commun à la Libye et au Tchad a notamment été mis en évidence par l’étude de l’anthracothère Libycosaurus (Lihoreau et al., 2006).
À la limite Miocène-Pliocène, la paléogéographie indique des relations complexes entre l’Asie, l’Afrique, et l’Europe avec des périodes de connexions et d'isolements (e.g. Benammi et al., 1996 ; Krijgsman et al., 1999b ; Gibert et al., 2013). La découverte en Afrique dans des gisements datés de la fin du Miocène supérieur ou début du Pliocène de taxons ayant un cachet européen a suscité un vif intérêt. La fin de la période Miocène est en effet critique pour la paléogéographie méditerranéenne car il se produit un événement sans précédent, la crise de salinité messinienne : la baisse du niveau de la mer a été tellement importante, plus d’un millier de mètres, qu’il est légitime d’y voir une source possible d’échange faunique entre le Maghreb et l’Europe sud-occidentale. Toutefois cette question s’est révélée très vite plus compliquée en raison de plusieurs facteurs. L’établissement du cachet d’un taxon qui suppose une détermination sûre, et la précision des datations et des corrélations sont indispensables pour fixer la position relative d’un site par rapport à la crise de salinité. Enfin, la période considérée est caractérisée par une activité tectonique considérable en Méditerranée, ce qui est susceptible d’avoir entrainé une paléogéographie variable et complexe à l’Ouest comme au centre du bassin, ayant permis ces échanges. L’étude des rongeurs permettra de reconsidérer les voies de dispersions de ces faunes mio-pliocènes et notamment de préciser leur âge et leur direction, entre les continents mais aussi au sein de ces continents.
En Afrique du Nord, les micromammifères sont si différents de ceux du nord de la Méditerranée que l’échelle biochronologique européenne n’est pas applicable. La zone de répartition de certaines espèces a changé au cours du temps en réponse à des variations climatiques et environnementales. Mais certaines espèces, en particulier de micromammifères, auraient été isolées
par diverses barrières (montagnes de l’Atlas, désert du Sahara dans le sud, les mers à l'Ouest et au Nord) et auraient développé des adaptations locales en réponse aux pressions environnementales (Stoetzel, 2013).
Les micromammifères fossiles provenant des gisements néogènes sont encore peu étudiés en Afrique du Nord, et les considérations taxonomiques rarement prises en compte. À ce jour, peu d’études ont été activement consacrées aux gisements à micromammifères d’Afrique du Nord Occidentale (Maghreb), mis à part quelques travaux majeurs de Jaeger (années 70-80), de Benammi (1993, 1997, 2001, 2006) et de Geraads (1982, 1998a) au Maroc, et d’Ameur-Chehbeur (1979, 1988) et Coiffait (1991) en Algérie.
Le présent travail s’inscrit de ce fait dans le cadre plus général de recherches et d’études de nouveaux gisements à micromammifères d'âge miocène terminal dans les dépôts continentaux d’Algérie et du Maroc. Le travail de terrain s’est déroulé durant les deux premières années de thèse. L’étude de ces nouveaux gisements a été effectuée dans le but d’identifier les espèces et de reconstituer les communautés et les paléoenvironnements correspondants. Les recherches effectuées sur le terrain ont permis d’augmenter significativement la connaissance des faunes du Miocène terminal des gisements étudiés et d’apporter de nouvelles précisions quant à leur datation. Les nouvelles faunes récoltées ont également permis d’établir la composition précise des communautés de micromammifères en Algérie et au Maroc durant le Miocène terminal.
L’objectif principal de cette thèse est de parfaire la connaissance des communautés mammaliennes de l’Algérie et du Maroc durant la fin du Miocène, ces dernières étant encore mal connues. Pour améliorer nos connaissances, plusieurs missions de terrains ont été organisées sur le terrain dans le bassin d’Aït Kandoula au Maroc, et les bassins de la Tafna et d’El Eulma en Algérie.
Pour mener à bien ce travail, une étude magnétostratigraphique est réalisée sur les deux bassins algériens (la Tafna et El Eulma). La magnétostratigraphie est une méthode précise pour tenter de résoudre des problèmes chronologiques. Cette étude a permis de bien dater les niveaux fossilifères intercalés dans les deux bassins algériens cités. Notons que l’étude
magnétostratigraphique des dépôts du Miocène supérieur du bassin d’Aït Kandoula a été déjà réalisée antérieurement dans le cadre d’un autre travail (Benammi et al., 1996).
Le deuxième objectif majeur de ce mémoire porte sur l’étude de la faune de rongeurs du bassin d’Aït Kandoula. C’est une région clé qui a joué un rôle important dans le cadre des échanges fauniques juste avant la crise de salinité messinienne. L’exploitation des trois nouvelles localités fossilifères de ce bassin a permis d’obtenir de nouvelles faunes de rongeurs à AF 12-1, AF12-2 et AF12-3. Ces faunes apportent une documentation considérable et inédite qui a fait l’objet d’une étude systématique approfondie. Celle-ci implique la comparaison exhaustive de ce matériel avec d’autres formes fossiles contemporaines et la proposition d’une histoire paléobiogéographique des rongeurs en intégrant les résultats précédents et en les confrontant aux données paléogéographiques et paléoclimatiques connues.
1
Introduction
Dans l’Ouest algérien, les bassins néogènes constituent un domaine complexe situé sur la marge méridionale de la Méditerranée, en avant d'une flexure cratonique (Perrodon, 1957). Au Sud, ces bassins sont bordés par les hautes plaines oranaises et au Nord par une série de massifs jurassiques alignés suivant l’axe Est-Ouest. En Algérie orientale, les terrains néogènes s’étendent de Sétif à Constantine. Ces bassins s’individualisent ainsi en deux termes : un terme marin dit Miocène « ante-nappe » débute soit avec le Miocène moyen soit avec le Miocène supérieur. Les formations continentales sont constituées principalement par des dépôts détritiques plus au moins grossiers associés à des marnes dans lesquelles s’intercalent parfois des niveaux carbonatés, évaporitiques, et volcaniques. La mise en place de ces bassins néogènes post-nappes a été accompagnée par une intense activité tectonique mio-pliocène (Guiraud, 1973).
Le Néogène continental au Maroc est essentiellement restreint au domaine atlasique et anti-atlasique. Ces dépôts se sont accumulés dans des bassins intra-montagneux ou dans des systèmes de vallées creusées dans des sédiments plus anciens, paléozoïques ou précambriens. Cette série continentale comporte des dépôts variés et hétérogènes constitués par des conglomérats, grès, marnes roses, et calcaires lacustres ou palustres. A l’Est du domaine atlasique, le Miocène continental est reconnu dans les Hauts plateaux par les calcaires anciennement attribués au Pliocène par Jodot (1955). Il est connu également sous forme de dépôts de comblement dans de grandes cuvettes synclinales comme celle de la plaine de Guercif-Taourirt. La présente étude s’est focalisée sur le bassin d’Aït Kandoula.
2
Localités et origines du matériel étudié
L’essentiel du matériel provient de différents bassins néogènes du Maroc (bassin d’Aït Kandoula) et d’Algérie (bassin de la Tafna et d’El Eulma) (Figure I. 1). Une étude magnétostratigraphique et paléontologique est menée sur les deux bassins algériens : El Eulma et Tafna. En revanche, l’étude paléontologique concerne essentiellement les dents de rongeurs récoltées
dans trois nouveaux gisements fossilifères d’Afoud (Maroc). Le présent travail, qui porte sur les rongeurs, a pour but de combler en partie les lacunes de connaissance sur les rongeurs et met en évidence l’intérêt qu’il peut y avoir à reprendre l’étude de ces taxons délaissés depuis longtemps.
Le contexte géologique des différentes localités sera détaillé dans les différents chapitres (ou articles et projets d’articles) qui suivent.
Figure I.1. Situation géographique des différents bassins étudiés dans le cadre de ce travail : 1) bassin
d’el Eulma, 2) bassin de la Tafna, 3) bassin d’Aït Kandoula.
3
Prélèvement du matériel
Par leur petite taille, les restes de micromammifères passent souvent inaperçus dans les dépôts. Seul un tamisage sous l’eau utilisant un tamis aux mailles voisines de 0,5 mm à 0,8 mm permet de les recueillir. Leur abondance relative varie avec le type de gisement. Dans les dépôts fluvio-lacustres et les complexes pédologiques, les restes provenant d’animaux ayant péri de mort naturelle sont souvent dispersés et rares. Dans les remplissages de grottes et les abris sous roches,
les restes de micromammifères qui proviennent de l’accumulation d’anciennes pelotes de rejections d’oiseaux rapaces sont souvent très abondants.
Le tamisage de quelques centaines de kilogrammes de sédiments ou la dissolution à l’acide de la gangue calcaire permet de recueillir une abondante microfaune. Après séchage, les résidus sont minutieusement triés avec une loupe binoculaire. Les dents et les restes osseux de micromammifères sont récupérés, puis classés par zone et par couche afin de connaitre leur origine exacte dans le gisement.
3.1
Mesures
Les mesures ont été prises sur l’ensemble des spécimens fossiles référencés dans cette étude. Le matériel dentaire comprend des prémolaires et des molaires. Les mesures sont exprimées en millimètres. Elles ont été effectuées à l’aide d’un mesuroscope monoculaire Mitutoyo associé à un compteur digital. Ces mesures correspondent aux diamètres mésio-distal maximal (L) et labio- lingual maximal (l) des couronnes des dents.
3.2
Taxonomie paléontologique
La détermination des différentes espèces de micromammifères est basée sur l’étude des premières molaires pour certaines familles (muridés et cricetidés), l’absence ou la présence de certains caractères dentaires pour certaines autres. Les données morphométriques ont également été prises en compte, notamment dans le cas d’espèces jumelles difficilement différenciables par des caractères morphologiques (par exemple Paraethomys meini et P. athmaniae).
3.3
Nomenclature
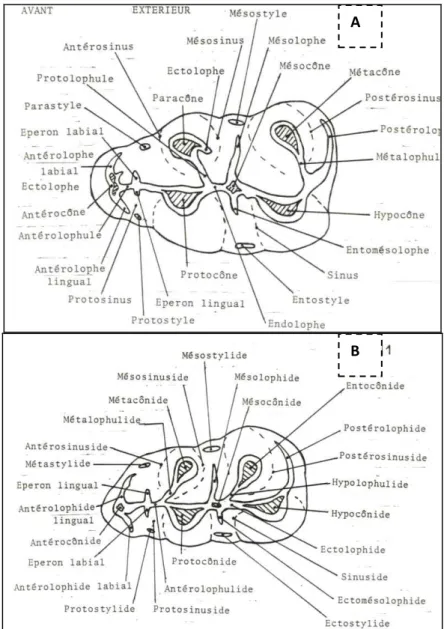
Les nomenclatures utilisées pour la description des caractères dentaires des micromammifères étudiés dans ce travail sont indiquées sur les schémas qui suivent. La nomenclature utilisée pour la description des dents des Myocricetodontinae et des Gerbillinae est la même que celle utilisée pour les Cricetidae (Figure I.1.1) (Mein et Freudenthal, 1971).
Figure I.1.1. Nomenclature des caractères dentaires des Cricetidae (d’après Mein et Freudenthal,
1971), en vue occlusale : A-molaire supérieure droite, B-molaire inférieure gauche
A
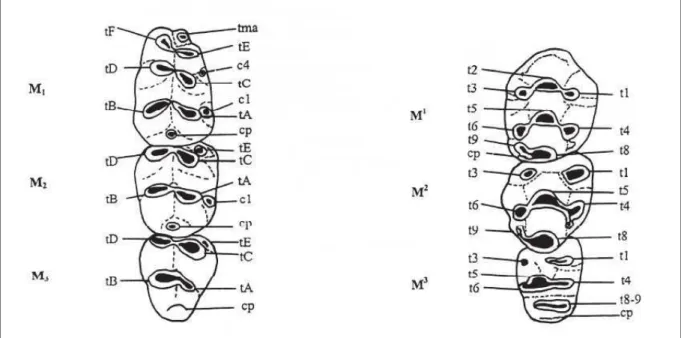
Figure I.1.2. Terminologie utilisée et méthode de mesures des dents de Muridae (d’après Michaux,
1971), rangée supérieure (à droite) et inférieure (à gauche). (t : tubercule, cp : cingulum postérieur).
Figure I.1.3 . Nomenclature dentaire des Gliridae et méthode de mesure : a) molaire supérieure
gauche. b) molaire inférieure gauche. c) méthode de mesure pour des dents isolées. L = longueur mésio-distale. L = largeur labio-linguale.
Figure I.1.4 . Nomenclature de la morphologie dentaire, et méthode de mesure des Sciuridae (van de
Weerd, 1976) : a) nomenclature dentaire des dents supérieures. b) Nomenclature dentaire des dents inférieures. c) Méthode de mesure pour des dents isolées.
Figure I.1.5. Nomenclature des caractères dentaires de Prolagus (Lagomorpha) : p3 : prémolaire
3.4
Photographie
Toutes les dents ont été photographiées au microscope électronique à balayage, avec des grossissements différents, selon la taille de la dent.
4
Biochronologie du Miocène continental en Afrique du Nord
L'étude des micromammifères fossiles est un outil fondamental pour établir une échelle biochronologique pour les dépôts continentaux, grâce à leurs évolutions rapides, leur abondance, et la facilité avec laquelle ils sont obtenus. L'élaboration d’une échelle basée sur des mammifères comporte néanmoins plusieurs difficultés : notamment, les conditions favorables pour la préservation de leurs restes ne sont pas fréquentes, et le registre fossile est très discontinu. Les gisements sont souvent des sites isolés sans relations géométriques claires entre eux ou séparés par quelques mètres de dépôts stériles. Ainsi, à la différence des dépôts marins, les limites entre les unités biostratigraphiques distinctes sont difficiles à établir.
En Europe, jusqu'aux années soixante, la terminologie employée pour établir l'âge des dépôts continentaux était basée sur l'échelle chronostratigraphique marine. Les gisements continentaux ayant livré un matériel peu abondant de mammifères étaient datés, quand c'était possible, par corrélation avec des sédiments marins intercalés dans ces sites fossilifères.
La première tentative de construction d’un cadre biostratigraphique pour les dépôts continentaux européens a été proposée par Thaler (1966), qui a divisé le Tertiaire en se basant sur des faunes de rongeurs, en assignant un niveau de référence à chacune d'elles. Cependant, certains de ces niveaux correspondent à des remplissages karstiques, qui ne permettent pas d’établir des corrélations lithostratigraphiques précises. Plus tard, Thaler (1972) a proposé une nouvelle échelle qui combine l’échelle de zones et l’échelle de niveaux repères, chaque biozone portant le nom d’un gisement qui en définit le niveau le plus élevé.
d’espèces caractéristiques est rapportée à chacune des zones. Mein (1975) a proposé des correspondances approximatives avec l’échelle marine. Une révision de la biozonation de Mein est parue en 1990 : les noms d’espèce y ont été abandonnés au profit de noms de genres, ce qui permet une meilleure corrélation entre des domaines géographiques éloignés. Les associations n’apparaissent plus. En revanche, les extinctions sont indiquées, et un gisement de référence est indiqué pour chaque zone ou sous-zone.
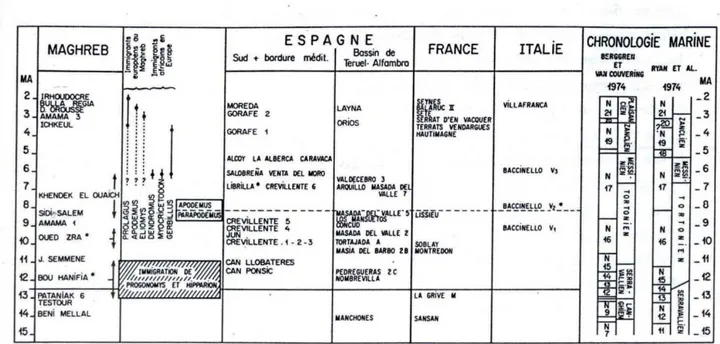
En Afrique du Nord, les taxons mio-pliocènes sont si différents de ceux d’Europe, que les échelles biochronologiques européennes ne sont pas applicables. Une échelle biochronologique d’Afrique du Nord a été réalisée par Jaeger et al. (1977) (Figure I.2.1). Ces auteurs ont proposé une biochronologie des gisements du Miocène supérieur d’Europe sud–occidentale et d’Afrique du Nord ainsi que des corrélations avec l’échelle stratigraphique marine. Ils regroupent ainsi les diverses faunes du Miocène supérieur d’Afrique du Nord en trois unités majeures :
- Le Mellalien : défini d’après les niveaux repères de Beni Mellal et Pataniak 6. Il est caractérisé par une faune qui se distingue par la présence de Myocricetodontinae dont Mellalomys atlasi et Myocricetodon parvus. Le genre d’Equidae Hipparion n’est pas encore représenté.
- Le Vallésien : défini d’après les niveaux repère de Bou Hanifia 1 et 2 et Oued Zra. Il est caractérisé par la présence simultanée du Cricetidae Zramys, du muridé le plus primitif en Afrique du Nord Progonomys cathalai, et d’une espèce primitive d’équidé : Cormohipparion primigenium.
- Le Turolien : défini d’après les niveaux repère d’Amama 2 et Khendek el Ouaich. Il est caractérisé par une association de faune plus évoluée que celles contenues dans les faunes précédentes (e. g. Paraethomys miocaenicus, Myocricetodon ouaichi).
De nombreux sites ont été découverts et publiés par plusieurs auteurs (e. g. Ameur-Chehbeur, 1979 ; Coiffait, 1991 ; Benammi et al., 1995 ; Benammi, 1997). La découverte de nouveaux taxons a amené des précisions importantes sur les lignées évolutives déjà connues, et a permis de mettre en évidence l’existence de nouvelles lignées.
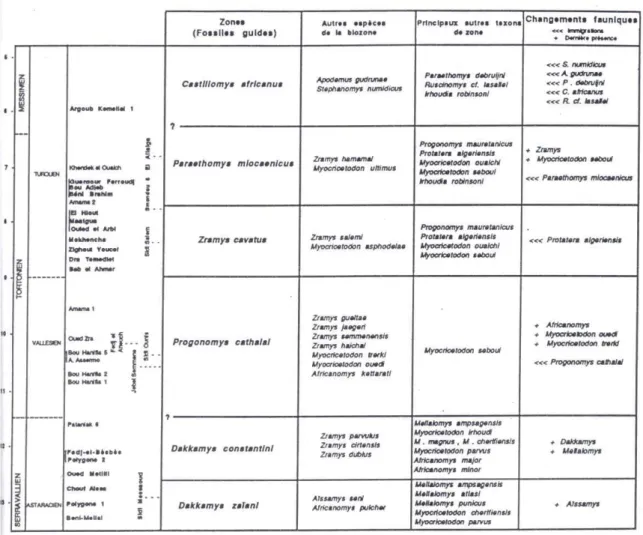
Coiffait (1991) a identifié six zones pour la période du Miocène moyen et supérieur. Ces zones sont fondées sur des «fossiles guides» (espèces de rongeurs caractéristiques) (Figure I.2.2). Les premières apparitions (FAD) et les extinctions (LAD) sont signalées, plutôt que d’employer des lettres ou des chiffres pour nommer ces zones.
Dans un travail plus récent, Stoetzel (2013) a proposé une nouvelle biozonation. Cette échelle biochronologique est une compilation des données antérieures, basée sur les premières apparitions des espèces (Figure I.2.3).
Figure I.2.1. Corrélation entre les formations continentales et marines dans le Néogène supérieur de
5
Magnétostratigraphie
La magnétostratigraphie est l’étude de la mémoire magnétique des roches et de sa succession temporelle. Lors de la mise en place des sédiments ou lors du refroidissement d’une roche volcanique, certains minéraux magnétiques acquièrent les caractéristiques du champ géomagnétique ambiant. L’analyse de la mémoire de ces roches permet de retrouver ce signal originel. Il est le témoin unique du champ géomagnétique et de la polarité de la dynamo terrestre à un moment de son histoire.
Au cours des temps géologiques, le champ magnétique de la Terre a connu une succession de polarités normales et inverses, de durée plus ou moins longue. L’échelle des inversions géomagnétiques est établie sur cette succession. La chronologie est basée sur des phénomènes géologiques ou biologiques. Pour que cette méthode puisse constituer un outil stratigraphique, il faut établir la stratigraphie magnétique d’une section avec la plus grande précision possible afin de la comparer avec l’échelle des inversions géomagnétiques. Le succès d’une étude magnétostratigraphique dépend à la fois des particularités de la coupe étudiée (aimantation des sédiments, taux de sédimentation, épaisseur, données chronologiques) et de la période concernée par la coupe.
Cette méthode a un vaste champ d’application. À présent, il est possible d’étudier la magnétostratigraphie de toutes les séquences sédimentaires, au fond des océans comme sur les terres émergées. Par ailleurs, les dépôts continentaux sont discontinus : les formations, particulièrement celles mises en place dans des formations fluviatiles, présentent souvent des lacunes de sédimentation. Par conséquent, dans ce type de dépôts continentaux, l’enregistrement paléomagnétique est moins constant que celui des fonds marins et les dépôts continentaux sont donc moins appropriés à ce genre d’étude. De même, les taux de sédimentation sont variables et difficiles à estimer. Par exemple, la même épaisseur de sédiments prélevée dans deux bassins continentaux différents ne représente que très rarement le même intervalle de temps et les taux de sédimentation dépendent de nombreux facteurs difficilement évaluables. Les roches sédimentaires sont souvent caractérisées par une aimantation faible et la qualité du signal paléomagnétique est moins servante
par rapport aux roches volcaniques. La magnétostratigraphie est un outil précieux pour tenter de résoudre des problèmes chronologiques, car elle permet de caler des séries biostratigraphiques indépendantes les une des autres, comme les faunes ou flores marines et continentales. Cependant, un calage est nécessaire pour l’interprétation du signal magnétique. La plupart des études magnétostratigraphiques sont accompagnées d’une étude de la zonation biostratigraphique. La précision de cette zonation dépend évidemment des espèces biologiques trouvées et de l’environnement sédimentaire.
5.1
Prélèvement et préparation des échantillons
5.1.1 Échantillonnage
Sur le terrain, l’échantillonnage consiste à prélever plusieurs échantillons orientés sur un affleurement ou site géologique sous forme de carottes ou blocs orientés. Des carottes de 25 mm de diamètre sont prélevées à l’aide d’une foreuse à essence (Figure I.3.1). La foreuse est équipée d’un foret diamanté amagnétique et d’un système de refroidissement à eau. L’orientation de la carotte s’effectue à l’aide d’un outil spécial qui s’insère autour de la carotte, qui est équipé d’une boussole magnétique, d’une boussole solaire, et d’un inclinomètre. La boussole solaire permet de vérifier la bonne orientation magnétique lorsque la météo est clémente. Avant d’être cassées, toutes les carottes sont orientées selon l’angle de plongement de la carotte par rapport à la verticale et l’azimut de son axe (Figure I.3.2). Ces valeurs sont ensuite notées sur le carnet de terrain. Sur la carotte retirée, l’axe de son azimut est gravé à l’aide d’un traceur.Chaque carotte est ensuite découpée en échantillons de taille standard de 22 mm de long. Selon la lithologie des faciès des niveaux échantillonnés, deux techniques de prélèvement sont déterminées par la qualité des affleurements. Pour le prélèvement des niveaux moins consolidés (e.g. argiles) qui ne peuvent pas être forés à l’eau, une autre méthode d’échantillonnage est appliquée. Elle consiste à prélever des blocs en place. On trace la direction du nord magnétique sur la surface structurale du banc à l’aide d’une boussole munie d’un niveau à bulle. Au laboratoire, les blocs sont inclus dans un support de plâtre. Ensuite, les blocs sont forés à l’aide d'air comprimé, perpendiculairement au plan orienté. Par la suite, les échantillons prélevés sont découpés au laboratoire en spécimens de 25 mm de long.
Figure I.3.1 . Méthode de prélèvement sur les bancs indurés : a) forage in situ ; b) orientation dans
l’espace ; c) trace de la génératrice.
Figure I.3.2. Orientation des carottes échantillonnées. Les coordonnées de la carotte sont définies à
partir de la génératrice. L’axe X est vers le haut, Y est perpendiculaire à X dans le même plan horizontal et Z est selon l’axe de la carotte dans le sens du forage (d’après Butler, 1992).
6
Les différents modes d’acquisition de l’aimantation dans les roches
sédimentaires
6.1
Acquisition de l’aimantation rémanente (ARN)
La plupart des roches sédimentaires enregistrent le champ magnétique ancien de différentes manières, présentant une aimantation rémanente naturelle. L’ARN est une aimantation rémanente
d'une roche avant tout traitement au laboratoire. Elle est généralement le produit de l’aimantation rémanente primaire et des aimantations rémanentes secondaires. Les roches peuvent acquérir leur aimantation par les processus suivants.
6.1.1 L’Aimantation Thermorémanente (ATR)
Cette aimantation est acquise par refroidissement en présence d’un champ magnétique, depuis une température supérieure à la température de Curie jusqu’à la température ambiante. L’ATR est parallèle et de même sens que le champ et elle est relativement stable dans le temps.
6.1.2 L’Aimantation Rémanente Isotherme (ARI)
Cette aimantation résulte de l’exposition à un champ magnétique fort (en comparaison au champ magnétique terrestre) pendant un temps court (quelques secondes à quelques minutes). Dans la nature, les courants électriques produits par la foudre ont le même effet. Dans le temps, l’ARI est moins stable que l’ATR.
6.1.3 L’Aimantation Rémanente Chimique (ARC)
Lorsqu’une roche est soumise à des processus de diagenèse, de métamorphisme, ou d’altération, de nouveaux minéraux ferromagnétiques se forment. Ils acquièrent une aimantation, qui est dite aimantation rémanente chimique (ARC). Les grains ferromagnétiques créés s’aimantent parallèlement au champ extérieur.
6.1.4 L’Aimantation Rémanente Visqueuse (ARV)
Cette aimantation correspond à une aimantation secondaire acquise par l’action du champ magnétique terrestre actuel sur la roche, à température ambiante. La caractéristique de cette aimantation est d’être instable dans le temps et détruite à faible température. D’un point de vue paléomagnétique, cette aimantation est considérée comme un bruit indésirable.
La plupart des roches sédimentaires présentent une aimantation naturelle. Cette aimantation est acquise lors de la mise en place des sédiments dans le champ magnétique terrestre ambiant. Au moment du dépôt, les particules magnétiques s’orientent parallèlement au champ magnétique existant et vont fossiliser la direction de ce champ (Figure I.3.3). La compaction ultérieure du sédiment produit souvent une erreur d’inclinaison.
Figure I.3.3. Schéma représentant l’acquisition de l’aimantation de type détritique lors de la
sédimentation de particules magnétiques. Le moment magnétique des minéraux magnétiques s’oriente le long des lignes de champ (d’après Tauxe, 2002).
6.2
Minéralogie magnétique
Pour l’étude paléomagnétique des roches, il est important de savoir quel est le minéral, ou les minéraux, porteur(s) de l’aimantation. L’identification du minéral magnétique dominant donne une idée sur le temps et le mode d’acquisition de l’ARN. Dans les roches sédimentaires, les minéraux magnétiques appartiennent à deux familles d’oxydes : les titanomagnétites et les sulfures. L’ARD due à la magnétite est considérée comme primaire. En revanche, les titanohématites peuvent être d’origine primaire ou secondaire. Dans ce dernier cas, il s’agit souvent d’un produit d’altération des titanomagnétites. D’autres minéraux magnétiques, généralement d’origine secondaire, peuvent également contribuer à l’aimantation rémanente du sédiment.
6.3
Méthodes d’identification des minéraux ferromagnétiques (Méthode
de Lowrie, 1990)
6.3.1 L’acquisition de l’aimantation rémanente isotherme (ARI)
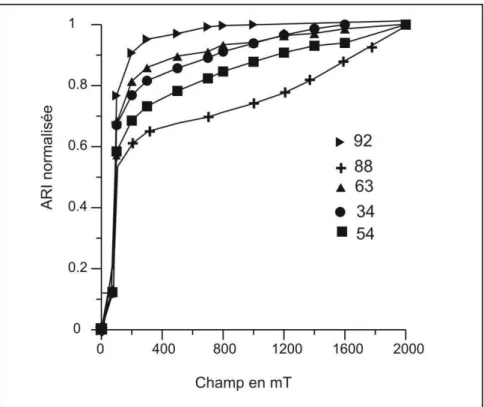
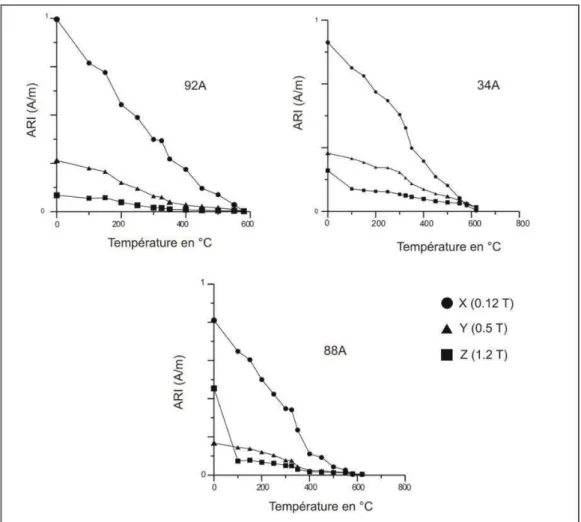
L’avantage de cette méthode, développée par Lowrie (1990), est qu’elle est basée sur la reconnaissance des minéraux ferromagnétiques à la fois d’après leur température de déblocage et d’après leur coercivité. L’acquisition des courbes d’aimantation rémanente isotherme (ARI) permet de différencier les minéraux en fonction de leur coercivité. Le principe en est simple. Léchantillon est soumis à trois champs différents dans trois axes orthogonaux. Le premier champ doit être suffisant pour saturer tous les minéraux ferromagnétiques et dans cette étude il est égal à 1,1 T appliqué selon l’axe Z de la carotte. L’ensemble des minéraux ferromagnétiques acquiert une aimantation rémanente isotherme dans cette direction. Le second champ (0,5 T) est ensuite appliqué selon l’axe Y et doit être suffisant pour saturer la magnétite mais pas les minéraux de forte coercivité comme la goethite, la pyrrhotite ou l’hématite. Le troisième champ (0,1 T) est appliqué selon l’axe X, et va remobiliser les minéraux de faible coercivité. Enfin, L’ensemble est désaimanté thermiquement. L’aimantation est mesurée sur chacun des axes à chaque étape de chauffe afin de déterminer le spectre de température de déblocage sur chacune des composantes.
6.3.2 Mesure de l’aimantation rémanente naturelle (ARN)
L’aimantation rémanente naturelle est l’aimantation acquise par les grains ferromagnétiques des roches au cours de leur histoire. Les directions de l’ARN ne reflètent qu’exceptionnellement la succession des polarités magnétiques sur une colonne stratigraphique. Il faut donc nettoyer l’ARN pour isoler ses composantes et identifier les directions de l’aimantation primaire. La désaimantation progressive de l’ARN permet de suivre l’évolution des directions de l’aimantation rémanente. Les échantillons sont chauffés progressivement jusqu’à la disparition totale de l’aimantation rémanente naturelle. Des chauffages par paliers jusqu’à 600°C-700°C mènent à une désaimantation progressive de l’échantillon et permettent de séparer les composantes d’aimantation portées par différents minéraux. L’aimantation rémanente est mesurée après chaque étape de chauffe à l’aide d’un
magnétomètre de type Spinner JR6 de marque AGICO. Les mesures de l’aimantation rémanente sont effectuées dans une pièce amagnétique.
•
Désaimantation
Les désaimantations thermiques et par champ alternatif permettent d’obtenir des renseignements sur les minéraux porteurs de l’aimantation rémanente. En effet, les températures de déblocage et les champs coercitifs sont des caractéristiques des différents minéraux ferromagnétiques.
La désaimantation par champ alternatif demande l’application d’un champ magnétique alternatif unidirectionnel de plus en plus fort (jusqu’à 100-150 mT) par paliers successifs. Cette méthode permet de débloquer tous les grains ayant une coercivité inférieure au champ maximum appliqué. La désaimantation par champ alternatif a l’avantage de préserver la minéralogie des échantillons, mais elle est inefficace pour supprimer la composante secondaire lorsque le sédiment comporte un minéral magnétique de haute coercivité résistant à ces traitements tels que l’hématite ou la goethite. Dans certains cas, la désaimantation des échantillons est ensuite poursuivie par des paliers de chauffe.
La désaimantation thermique est réalisée dans un four à blindage amagnétique d’une capacité maximale de 80 échantillons et dans lequel la température peut atteindre 700°C. Le traitement thermique commence en général à 100°C, puis la température est augmentée par pas de 25 °C à 50 °C au cours des étapes de désaimantation. Dans certains cas, la désaimantation thermique permet d’identifier les composantes de l’ARN de manière plus efficace que la méthode précédente tout dépend de la nature des minéraux.
7
Traitement des données
7.1
Acquisition et visualisation des données
Les résultats des désaimantations thermiques et par champ alternatif sont représentés dans des diagrammes de Zijderveld (1967). Il s’agit d’une projection classique en géométrie descriptive. On projette la trace de l’extrémité du vecteur aimantation sur deux ou trois plans orthogonaux. Le plan horizontal le long duquel sont projetés la déclinaison et un plan vertical qui permet de définir l’inclinaison. Sur cette projection, il est facile de voir si l’aimantation change ou non de direction, si elle décroit ou non, et d’en séparer différentes composantes.
Lorsqu’une aimantation est portée par une seule composante, l’aimantation se détruit toute seule, et les points représentatifs des paliers de désaimantation se placent sur une droite. Dans d’autres cas, où l’aimantation est portée par plusieurs composantes, si les porteurs de l’aimantation ont des champs coercitifs ou des températures de blocage bien distincts, on obtient une ligne brisée dont chaque segment correspond à des aimantations présentes dans l’échantillon. L’aimantation primaire est généralement plus résistante. Ainsi, il est facile de voir si l’aimantation change de direction, si elle décroît ou non, et de séparer aussi les différentes composantes. La droite moyenne qui traverse le groupe de données pour la composante de l’ARN est examinée grâce à l’analyse en composantes principales (Kirschvink, 1980). Les segments identifiés possèdent N points de données, cette méthode consiste à calculer son « centre de masse »:
La direction moyenne est calculée par la méthode de Fisher (1953) pour chaque site défini. Les différents paramètres statistiques permettent d’avoir des incertitudes sur la direction moyenne calculée. Si les directions d’aimantation suivent une loi de Fisher, adaptation à la sphère de la loi de Gauss ou la loi normale sur un plan (Fisher, 1953), le rayon du cône de confiance à 95 % autour de la moyenne est calculé. Il est noté α95 dans les données. La distribution est dite « fisherienne » lorsque la distribution présente une symétrie de révolution autour de la direction moyenne. Le paramètre de groupement k des mesures est calculé : si k est grand, la distribution sera très regroupée ; si k est petit la répartition sera étalée. Les directions moyennes obtenues pour chaque site sont transformées
directions obtenues à partir des paléopôles pour dater l’acquisition de l’aimantation rémanente naturelle.
7.2
Tests paléomagnétiques
Afin d’évaluer si les directions d’aimantation peuvent être considérées comme primaires, secondaires ou aléatoires, il existe plusieurs tests :
Test du pli
Ce test permet de déterminer si l’âge de l’aimantation est antérieur, postérieur ou synchrone par rapport à la déformation. Afin de réaliser ce test, plusieurs échantillons sont prélevés sur les deux flancs opposés d’un pli. Les directions de l’aimantation primaire in situ (sans correction), puis les directions avec correction stratigraphique, sont calculées. Un test de pli est dit positif si le meilleur groupement des directions d’aimantation des échantillons provenant de deux flancs d'un pli est obtenu après correction de pendage. Dans ce cas, l'aimantation est acquise avant la déformation (l’acquisition de l’aimantation est antérieure à l’âge du plissement), impliquant la nature primaire de l’aimantation. En revanche, si les directions sont mieux groupées avant la correction, l'aimantation est postérieure au plissement, ce qui constitue un test de pli négatif (Figure I.3.3).
Figure I.3.3. Exemple de test de pli. Le graphique de gauche montre les directions avant la correction
du pendage de deux groupes de directions échantillonnées sur les deux flancs d’un pli. Sur le graphique de droite, les mêmes directions, représentées après correction de pendage, montrent un meilleur groupement ce qui donne un test de pli positif (d’après Tauxe, 2002).
• Test d'inversion
Le champ magnétique a subi des inversions au cours des temps géologiques. Le test de McFadden et McElhinny (1990) permet d’évaluer l’antipodalité de deux directions. Les directions d’aimantation sont considérées comme quasiment constantes avant et après une inversion. Néanmoins, les vecteurs d’aimantation de polarité opposée acquis dans des niveaux voisins doivent être antiparallèles. Un non antiparallélisme des deux directions montre qu’une composante secondaire s'est ajoutée à la composante principale. Un test d’inversion positif permet d’argumenter une nature primaire de l’aimantation.
Partie 1 : Etude magnétostratigraphique et
paléontologique des sites continentaux néogènes
Chapitre II. Etude magnétostratigraphique et
biochronologique du Bassin d’El Eulma
1
Introduction
Jaeger (1977a) a attiré l’attention sur le gisement d’Amama en étudiant les rongeurs des trois gisements fossilifères (Amama 1, Amama 2 et Amama 3). Par la suite, d’autres travaux sont venus s’ajouter pour enrichir nos connaissances sur les faunes de rongeurs de ce bassin, avec une datation radiométrique qui a permis d’établir une échelle de corrélation entre les différents bassins continentaux de l’Afrique du Nord (Jaeger, 1977a ; Ameur-Chehbeur, 1988 ; Coiffait, 1991). L’âge des faunes de micromammifères découvertes dans les dépôts continentaux du bassin néogène d’El Eulma s’étend du Miocène supérieur au Pliocène moyen. Les trois niveaux fossilifères se succèdent le long d’une section d’environ 140 m d’épaisseur.
Lors de nos prospections géologiques menées sur ce bassin, nous avons choisi une nouvelle coupe affleurant au niveau d’un talweg se situant à 300 m plus au Nord par rapport à la coupe classique décrite par les anciens auteurs (Guiraud, 1973 ; Jaeger, 1975 ; Ameur-Chehbeur, 1988). Cette coupe présente trois niveaux cinéritiques dont un seul a été daté de 7.7 Ma ± 0.7 Ma (Coiffait, 1991). Dans cette nouvelle coupe se succèdent également trois niveaux fossilifères (Amama 1, Amama 2 et Amama 3), dont les faunes sont bien déterminées. Ce travail présente également les résultats de l’étude magnétostratigraphique de la coupe de Koudiat el Amama, dont le but est d’établir une corrélation avec l’échelle de polarités géomagnétiques afin d’attribuer des âges numériques pour ces trois niveaux fossilifères.
2
Cadre géographique et géologique
A l’Est de Sétif et au Nord d’El Eulma (Ex Arnaud), l’érosion régressive de l’Oued Deheb et de ses affluents a entamé les carapaces calcaires quaternaires qui recouvrent généralement les hautes plaines constantinoises, faisant ainsi apparaitre un petit bassin continental mio-pliocène conservé dans une cuvette entourée par les formations allochtones du Tell sétifien (Figure II.1). Le Bassin d’El
(Guiraud, 1973) du Miocène supérieur, conservé dans une cuvette, et se trouvant entouré au Nord, à l’Est et à l’Ouest par des unités telliennes allochtones (nappe de Djamila), tandis qu’au Sud il s’étale sur les hautes plaines sétifiennes. C’est un bassin bien individualisé par des dépôts d’une épaisse série dont l’âge s’étend du Miocène inférieur au Pliocène supérieur. Guiraud (1973), pour établir des corrélations avec les séries d’El Hodna, a donné une description des termes supérieurs dans la région de la Koudiat el Amama. Par la suite, Jaeger (1977a) a également fourni une coupe géologique de ce secteur (Figure II.2).
Figure II.1. Localisation géographique et géologique du Bassin d’El Eulma (Modifié d’après Sahnouni
D’après cet auteur, les couches les plus anciennes seraient visibles sur la partie septentrionale du bassin et seraient d’âge crétacé ou oligocène. Le Néogène du Bassin d’El Eulma se compose des termes suivants :
1) à la base, de puissants dépôts détritiques rouges reposant en discordance sur les terrains crétacés et éocènes des unités telliennes. Ces conglomérats sont épais de plusieurs dizaines de mètres et passent progressivement vers le haut à :
2) des marnes sableuses grises à rosâtres (à passées détritiques peu abondantes) développées également sur plusieurs centaines de mètres.
3) dans le secteur de Koudiat el Amama, une soixantaine de mètres de gypse en bancs métriques, séparés par des inter-lits d’argile grise. Cette passée de gypse semble latéralement se rapprocher à des calcaires lacustres et des marnes noires.
4) des argiles grises, présentant quelques passées gréseuses en lentilles plus ou moins importantes, riches en bivalves Unionidae, alors que les argiles grises encaissantes contiennent plutôt des reste de gastéropodes Helicidae. L’épaisseur visible de ce niveau est de 80 m environ.
5) recouvrant en faible discordance angulaire les formations précédentes, des limons rougeâtres, représentant la base des formations pliocènes d’El Eulma.
Figure II.2. Coupe du Bassin mio-pliocène d’El Eulma (Algérie), avec une localisation des principaux
3
Description de la coupe
La nouvelle coupe, épaisse de 140 m, montre des variations lithologiques de faciès, ainsi que la présence d’une importante discontinuité sédimentaire matérialisée par une discordance angulaire, qui permet de distinguer deux formations qui sont de bas en haut (Figure II.2) :
3.1
Formation I
À Koudiat el Amama, cette formation représente une alternance de marnes grises où s’intercalent des calcaires lacustres et des passées gypseuses. À cet endroit, cette formation est caractérisée par des grands paquets de marnes noires (riches en gastéropodes), alternées avec quelques passées gréseuses. La fréquence de ces dernières augmente de bas en haut. Dans cet ensemble s’intercalent trois niveaux cinéritiques. Le premier niveau, de 0,4 m d’épaisseur, est mis en évidence pour la première fois, et il occupe la base de la formation. Les deux autres niveaux (successivement de 4 m et 0,8 m d’épaisseur) apparaissent vers la base du dernier tiers de la formation. Nous signalons également la présence pour la première fois d’un troisième niveau cinéritique. Les niveaux gréseux se distinguent par des stratifications obliques et passent latéralement à des chenaux. Ces deniers sont occupés par des microconglomérats qui contiennent parfois des restes de vertébrés.
Le matériel prélevé (lavé, trié et analysé) dans différents niveaux argileux ou microconglomératiques de cette formation a révélé la présence de restes de micromammifères dont la composition faunique est analogue à celle des deux sites (Amama 1 et Amama 2) décrits par les anciens auteurs (Jaeger, 1977a ; Ameur-Chehbeur, 1988 ; Coiffait, 1991) leur permettant de suggérer un âge miocène supérieur à cette localité.
Le milieu de dépôts de cette formation est caractéristique d’une sédimentation continentale de type fluviatile.
3.2
Formation II
Cette formation repose en discordance angulaire sur la formation précédente. Elle démarre par des conglomérats qui passent latéralement à des grès grossiers renfermant quelques fois des restes de vertébrés. D’un endroit à un autre, l’épaisseur de cet ensemble est variable (2 m à 5 m). Au dessus, reposent des argiles rouges gréseuses à concrétions carbonatées qui passent graduellement à des argiles grises et des calcaires lacustres friables (15 m environ). Un niveau marneux de couleur sombre situé vers le sommet de cette alternance a livré des gastéropodes continentaux, des débris de végétaux, des restes d’Hipparion (Equidae) et des micromammifères (Amama 3). Plus haut dans la séquence, le faciès change considérablement et passe à une épaisse série de marnes sableuses rouges (30 m environ) où s’alternent quelques fois de petits bancs (0.5 m) de calcaire lacustre qui disparaissent latéralement. Ces dépôts s’amincissent vers l’Ouest et le Sud du bassin où ils ont livré les célèbres faunes de vertébrés d’Aïn Boucherit et d’Aïn Hanech (Arambourg, 1969). Ces faunes sont composées surtout de grands mammifères associés à une industrie lithique plus ou moins abondante (Sahnouni et al., 1996).
De manière générale, la sédimentation de ce vaste complexe continental montre une évolution d’un environnement lagunaire à la base qui passe ensuite à un milieu lacustre puis à l’installation d’un régime sédimentaire fluvio-lacustre vers le sommet de la coupe.
4
Données biostratigraphiques
Les trois sites fossilifères superposés (Amama 1, Amama 2 et Amama 3) ont été découvert par Guiraud (1973) et Jaeger (1977a), puis Ameur-Chehbeur (1988) a réalisé une étude détaillée des rongeurs. Lors d’une prospection paléontologique en Mars 2012 nous avons échantillonné dans un thalweg parallèle à celui que les précédents auteurs avaient échantillonné (Jaeger, 1977a ; Ameur-Chehbeur, 1988). La nouvelle coupe que nous présentons dans ce travail (Figure II. 3) a également livré les mêmes niveaux fossilifères, avec la même composition faunique que les travaux antérieurs. Les trois niveaux à micromammifères signalés dans cette localité sont de bas en haut:
Amama 1 : Ce niveau inférieur est situé à une quarantaine de mètres au dessus du toit des passées
calcaires. Ce gisement a fourni la liste faunique suivante : Schizogalerix sp. (Erinaceidae), et les rongeurs Atlantoxerus sp., Zramys cf. haichai, Progonomys cf. cathalai et Irhoudia sp. (Jaeger, 1977a). Une autre prospection paléontologique a été effectuée par Ameur–Chehbeur (1988) dans le cadre de sa thèse, ajoutant à la liste faunique précédente deux espèces de rongeurs : Myocricetodon cf. seboui et M. cf. trerki. Le degré d’évolution de la population de Zramys cf. haichai est plus élevé que celui de la population de l’Oued Zra ; ceci suggère un âge légèrement postérieur à celui de ce dernier gisement. Comme celui-ci est attribué au Vallésien supérieur, il est donc logique d’attribuer au gisement d’Amama 1 un âge proche de la limite Vallésien-Turolien (Jaeger et al., 1973).
Amama 2 : Ce niveau est situé dans une lentille d’argile noirâtre riche en gastéropodes, située sous le
toit de grès à galets qui a livré des restes d’Hipparion sitifense (Equidae). Ce toit de grès est situé vers le sommet de la formation miocène. Ce gisement a fourni la liste de micromammifères suivante : Schizogalerix sp. (Erinaceidae), et les rongeurs Atlantoxerus sp., Zramys hamamai, Protatera algeriensis, Myocricetodon cf. seboui, Paraethomys cf. miocaenicus, et Irhoudia robinsoni (Jaeger,
1977a). De même, Ameur–Chehbeur (1991) a signalé un nouveau genre et nouvelle espèce de gerbillidé Eulmus miocaenicus.
Les Cricetidae sont représentés par trois formes distinctes alors que les Muridae, les Ctenodactylidae et les Sciuridae ne sont représentés que par une seule espèce. Sur le plan quantitatif toutefois les Gerbillidae représentés par Protatera algeriensis ainsi que les Ctenodactylidae, représentés par Irhoudia robinsoni dominent largement. Ces proportions font apparaitre la domination de formes caractéristiques, en Afrique du Nord, des milieux semi-arides, ce qui suggère l’existence d’un biotope relativement ouvert et sec.
La position dans l’échelle biochronologique de ce gisement, situé au sein de la formation exclusivement continentale, n’est pas aussi aisée à mettre en évidence que celle de Khendek el Ouaich. Sa position stratigraphique, est localisée à 50 m au dessus du gisement d’Amama 1. Les données biochronologiques plus précises peuvent être tirées de l’étude des degrés évolutifs. Zramys hamamai est considéré comme le descendant direct de Z. salemi du gisement turolien inférieur de Sidi Salem (Algérie). Myocricetodon cf. seboui et Paraethomys cf. miocaenicus peuvent être comparés respectivement avec M. ouaichi et P. miocaenicus du gisement du Khendek el Ouaich. Les degrés évolutifs de ces deux espèces du gisement d’Amama 2 sont plus primitifs que ceux correspondant aux espèces de Khendek el Ouaich. D’après ces données, il conviendrait d’assigner au gisement Amama 2 un âge intermédiaire entre celui du gisement Turolien inférieur de Sidi Salem (Algérie) et celui du gisement du Khendek el Ouaich qui est situé au sein des formations régressives du Miocène supérieur marocain. C’est donc un âge turolien moyen qui est attribué à ce gisement.
Amama3 : Ce gisement a été découvert pour la première fois par Guiraud en 1973, à la base des
formations rouges. Celles-ci ont été attribuées au Villafranchien par Arambourg (1959), mais pour Ginsburg (1957), elles correspondent au Pliocène et au Villafranchien. Ce gisement provient de la même série, à une vingtaine de mètres au dessus de la discordance le séparant du gisement d’Amama 2, où les minces intercalations de marnes et de calcaires travertineux font leur apparition. Il s’agit d’un niveau très riche en matière organique, qui a livré la liste faunique suivante : les rongeurs