TWEAK : un nouveau régulateur de la ribonucléase III Dicer
Texte intégral
Figure
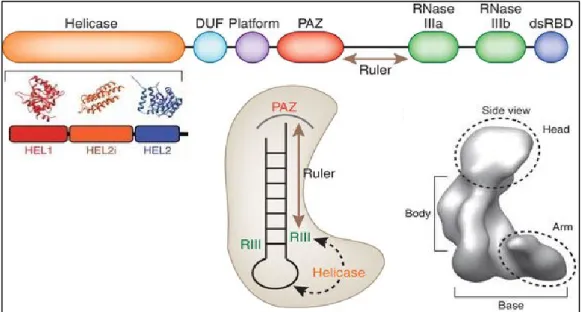
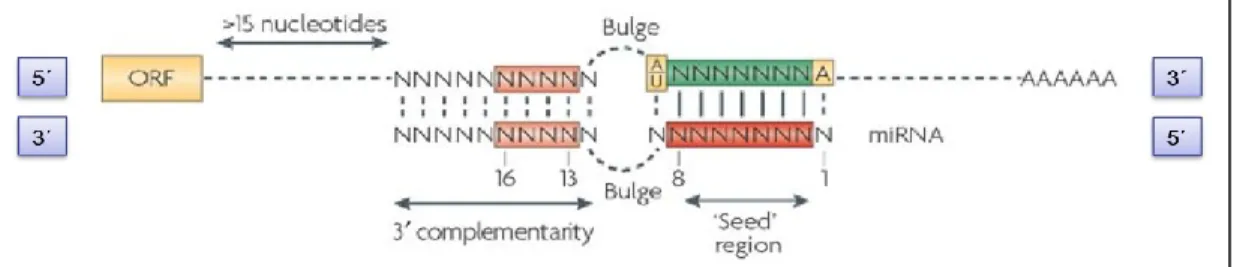




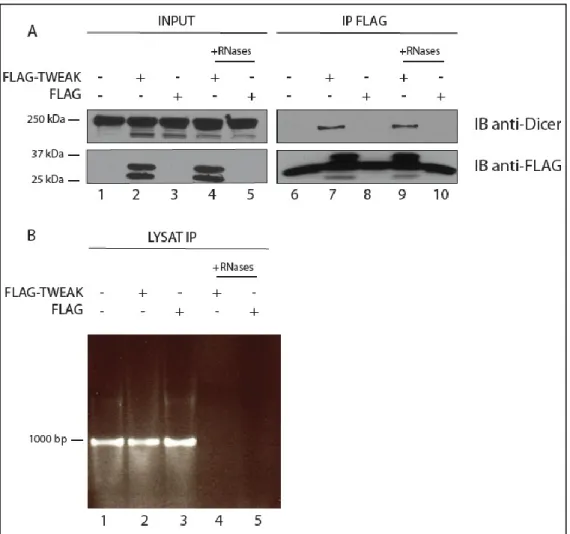


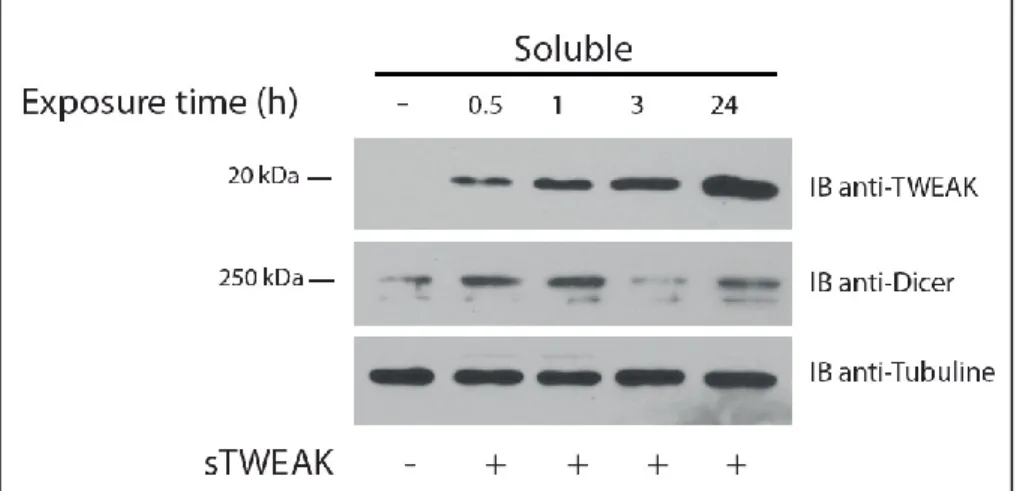
Documents relatifs
[r]
Although the comparison between the virulence of parasites in single infections and their overall viru- lence in multiple infections is relevant for a number of issues (e.g.
Visualization of ATP levels inside single living cells with fluorescence resonance energy transfer-based genetically encoded indicators.. An activity-induced microRNA controls
ةﺪﺤﺘﳌا تارﺎﻣﻹا 3 ( ءاﺮﺸﻟا ﰲ ﺔﻳﺰﻛﺮﳌا : ﻞــﻤﻌﺗ ﻦﻫﺪــﻠﻟ ﺔــﻴﻨﻃﻮﻟا ﺔــﺴﺳﺆﳌا ﺔــﺻﺎﺧ و ﺮــﺋاﺰﳉا ﰲ ﺔﻳدﺎــﺼﺘﻗﻻا تﺎــﺴﺳﺆﳌا ﻦــﻣ اﲑــﺜﻛ أﺪــﺒﲟ
The increase of total body weight included an increase in skeletal muscle mass, as demonstrated by signi ficant increase in type II-rich TA, EDL, and gastrocnemius muscle mass in
Breathing activity was described in terms of ventilatory flow measured using a PNT and, either simultaneously or sequentially, with a SONAR device measuring the velocity of the
results are used to identify periods when biomass burning was likely to have had a significant impact on the concentrations of elemental carbon at two sites, T1 and T2, downwind of
Adjusted all-cause mortality curves in asymptomatic and minimally symptomatic patients with severe aortic stenosis (n =559) according to mean transaortic pressure gradient
