Tomographie à rayons X; analyse et modélisation de l'ontogénèse des épicormiques du chêne sessile (Quercus petraea (L.) Matt.).
214
0
0
Texte intégral
Figure
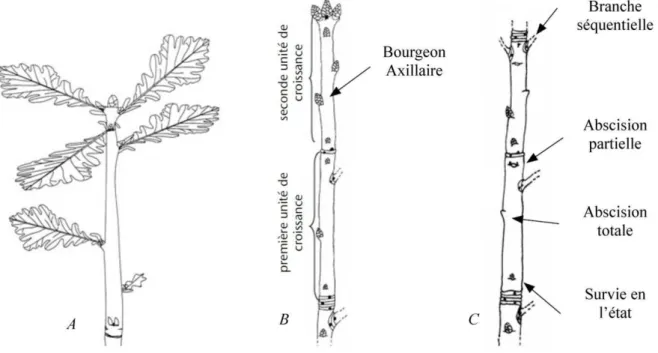
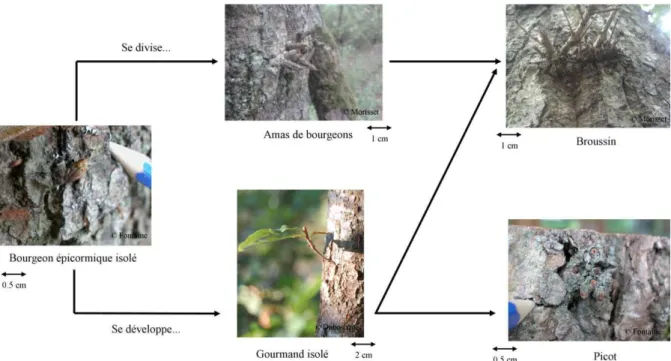

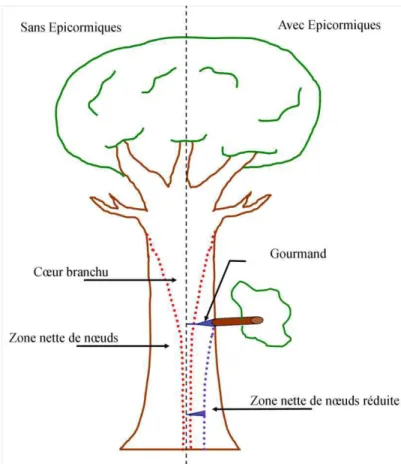
+7

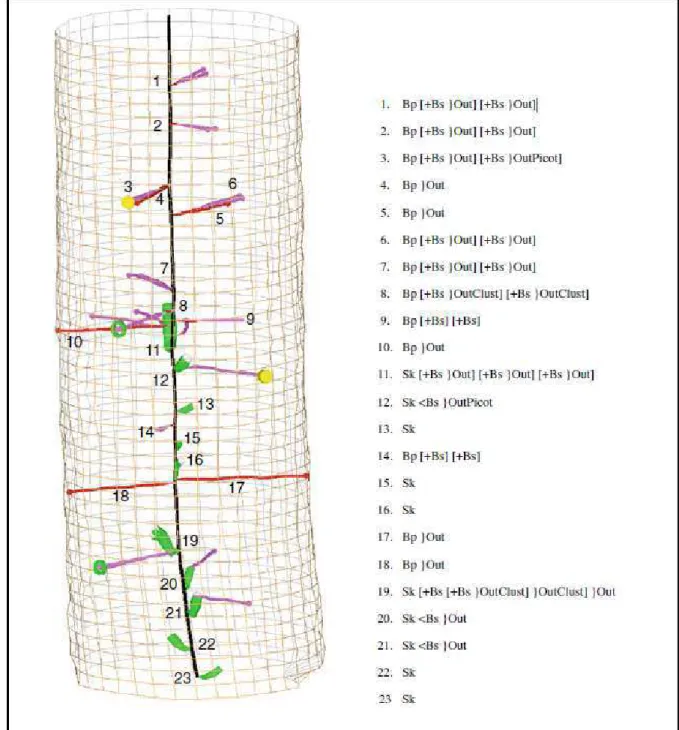
Documents relatifs
