Comment le dérèglement de la transcription peut être la cause d’avortements
Texte intégral
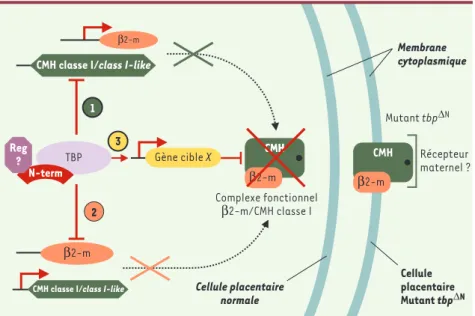
Figure
Documents relatifs
L’AMIU a été la méthode la plus utilisée dans les avortements incomplets et les œufs clairs avec respectivement 91,8% et 75% alors que le traitement médical avec le Misoprostol
Les membres de la communauté épistémique dont il est question ici sont inspirés par cette nouvelle identité mais sont à la fois plus liés par des causal beliefs (Haas,
Tableau 3. Toutes Ies données sont fondées sur les certificatç de décès, sauf pour I'Angleterre et Ie Pays de Galles OU les d&&s sont enregistrés sur
Ajoutons également que si les actes directs de discrimination sont moins importants dans le milieu éducatif que dans le milieu professionnel notamment (Luciak,
Yvonne, sa mère, Milou, sa sœur, et Simone se retrouvent dans la file destinée à intégrer le camp d’Auschwitz-Birkenau.. Elles sont ensemble et ne se
En 1993, le lien entre ces deux méca- nismes fondamentaux que sont la transcription et la réparation était enfin conforté ; nous montrions le rôle indispensable de TFIIH, défini
• Souvent, l’avortement se produit après des semaines voire des mois de l’infection initiale et son titre d’anticorps est stable ou en diminution au moment de l’avortement.
Devant l’absence de statistiques nationales récentes, nous avons conduit cette étude à la maternité du centre hospitalier de Libreville (CHL) dans le but de faire l’état des
