HAL Id: hal-02816977
https://hal.inrae.fr/hal-02816977
Submitted on 6 Jun 2020HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Fonctionnement hydraulique de la feuille de peuplier et
implication des aquaporines
David López
To cite this version:
David López. Fonctionnement hydraulique de la feuille de peuplier et implication des aquaporines. [Stage] Université Blaise Pascal (Clermont Ferrand 2) (UBP), FRA.; Ecole Nationale d’Ingénieurs des Travaux Agricoles de Clermont-Ferrand (ENITAC), FRA. 2009, 36 p. + annexes. �hal-02816977�
MASTER II Biologie et Environnement
SPECIALITÉ : GEPV
RAPPORT DE STAGE PRESENTE PAR:
DAVID LOPEZ
SUJET :
Fonctionnement hydraulique de la feuille de peuplier et
implication des aquaporines
RESPONSABLE DU STAGE
Aurélie Gousset
Juin 2009
UMR Physique et Physiologie intégratives de l’arbre fruitier et forestier
de diffusion de l’eau et du dioxyde de carbone chez les animaux et les plantes.
Le génome de Populus trichocarpa contient 63 gènes codant des aquaporines putatives. 16 sont homologues aux PIP d’Arabidopsis thaliana, 20 aux TIP, 13 aux NIP, 7 aux SIP, et 7 se distinguent des familles de MIP décrites jusqu’à présent. Une analyse des profils d’expression des PIP dans les différents organes montre une expression différentielle de certaines isoformes.
Des mesures de conductivité hydraulique foliaire chez deux espèces de peuplier (P.
nigra et P. trichocarpa) montrent une augmentation en réponse à la lumière. Cette réponse
peut être modulée par l’âge des feuilles ou une acclimatation à l’obscurité de celles-ci. En plus des expériences d’écophysiologie, une approche de biologie moléculaire a été menée pour caractériser la réponse à la lumière. L’étude de l’expression des gènes codant pour des PIP1 et PIP2 par qPCR, montre une régulation potentielle en réponse à la lumière d’une partie de ces gènes.
Ces résultats suggèrent que l’expression des PIP de peuplier est régulée par des facteurs endogènes et exogènes ce qui impacte potentiellement la capacité des organes concernés à être perméables à l’eau.
MOTS CLES
Peuplier, conductance hydraulique foliaire, lumière, aquaporines, qPCR.
ABSTRACT
Aquaporins are intrinsic membrane proteins known to influence the movement of small molecules including water and CO2 in animals and plants.
Populus trichocarpa genome contains 63 genes coding for putative aquaporins. 16
genes are homologous with Arabidopsis thaliana’s PIP, 20 belong to the TIP family, 13 to NIPs, 7 to SIPs, and 7 genes differ from the MIP family members described so far. An analysis of PIP-type genes expression patterns in different organs shows a differential expression for some of the isoforms.
Leaf hydraulic conductance measurements carried out on two poplar species (P. nigra and P. trichocarpa) shows an increase in leaf hydraulic conductance in response to light, which can be modulated by leaf age or acclimatization to dark. In addition with the ecophysiological experiments, a molecular biology approach was carried out for light response characterization. Real-time qPCR analyses of PIP1 and PIP2 family aquaporins coding genes shows a possible regulation in response to light for some of the genes.
These results suggest that poplar PIP expression is regulated by endogenous and exogenous factors, hence having a potential impact on concerned organs water transport capability.
KEY WORDS
A : assimilation nette de dioxyde de carbone
ADNc : acide désoxyribonucléique complémentaire ARN : acide ribonucléique
ATP : adénosine triphosphate bp : paires de base
CaCl2 : chlorure de calcium CDS : coding sequence
Ci : concentration en CO2 dans les espaces intercellulaires Cc : concentration en CO2 dans les chloroplastes
cm : centimètre
CO2 : dioxyde de carbone Ct : threshold cycle °C : degré Celsius
dNTP : désoxyribonucléotide EFM : evaporative flux method EST : expression sequences tags F : transpiration
gm: conductance mésophyllienne ou interne au CO2 H : heure
H2O : eau
H2O2 : peroxyde d’hydrogène HgCl2 : chlorure de mercure HPFM : high pressure flow meter Hv : Hordeum vulgare
Jmax : flux maximal d’électrons
K : conductance hydraulique ou conductivité hydraulique KCl : chlorure de potassium
kDa : kilodalton kPa : kilopascal M: mètre Min : minutes
MIP : major intrinsic protein MPa : mégapascal
NIP : nodulin26-like intrinsic protein O2 : dioxygène
OAA : oxaloacetate Pa : pascal
PAR : photosynthetically active radiation, radiations efficaces pour la photosynthèse PGA : 3-Phosphoglycerate
PIP : plasma-membrane intrinsic protein
PCR: polymerase chain reaction, réaction de polymérisation en chaine Pt : Populus trichocarpa
qPCR : quantitative PCR R : résistance hydraulique Rd : respiration mitochondriale RKM : rehydration kinetics method RT: reverse transcription
SIP : small basic intrinsic protein TIP : tonoplast intrinsic protein TM : segment transmembranaire Tm: température de fusion UTR: untranslated region
Vc : assimilation du CO2 par carboxylation du ribulose 1, 5 bisphosphate Vcmax : vitesse maximale de carboxylation
V0 : dégagement de CO2 par photorespiration W : watt
WUE: water use efficiency, efficience d’utilisation de l’eau Zm : Zea mays
BIBLIOGRAPHIE
I- Conductance hydraulique ... p1 1- Conductance hydraulique et résistance foliaire... p1 2- Sources de variations de la conductance hydraulique foliaire ... p2 II- Photosynthèse et conductance mésophyllienne au CO2... p2 1- Photosynthèse... p3 2- Conductance mésophyllienne au CO2... p4 III- Les aquaporines ... p5 1- Les différentes familles de MIP ... p6 a- La famille des PIP... p6 b- La famille des TIP ... p6 c- La famille des NIP... p7 d- La famille des SIP ... p7 2- Expression des aquaporines ... p7 3- Les aquaporines : à l’interface entre métabolisme carboné et fonctionnement hydraulique ? ... p8 IV- Le peuplier, modèle d’étude ... p8 V- Objectifs du stage... p9 TRAVAIL EXPERIMENTAL ... p11 MATERIEL ET METHODES
I- Matériel végétal ... p11 II- Approches d’écophysiologie ... p11 1-Mesures de photosynthèse... p11 a- Réponse au CO2... p11
b- Calcul des paramètres photosynthétiques... p12 2- Mesure de conductance hydraulique foliaire ... p12 a- Etude de variations en réponse à la lumière ... p12 b- Recherche d’une régulation endogène de la conductance hydraulique... p13 c- Etude du rôle de la qualité de la lumière ... p13 III- Approches de biologie moléculaire ... p13 1- Séquences des aquaporines ... p13 2- Clonage des aquaporines ... p13 3- PCR quantitative en temps réel ... p14 a- Amorces utilisées... p14 b- Extraction des ARN et reverse transcription... p 14 c- Matériel utilisé et programme PCR ... p15 d- Profils d’expression tissus spécifique des PIP1 et 2 chez Populus.
trichocarpa ... p 15
e- Profils d’expression en réponse à la lumière des PIP1 et 2 chez Populus
trichocarpa et Populus nigra... p15
4- Arbre phylogénétique... p16 RESULTATS
I- Caractérisation des aquaporines de Populus trichocarpa... p17 1- PIP1 ... p17
II- Caractérisation de deux espèces de peupliers : Populus trichocarpa et Populus nigra ... p18 1- Photosynthèse... p19 2- Conductance hydraulique foliaire ... p20 a- Age de la feuille... p20 b- Qualité de la lumière ... p 20 c- Rythme endogène ... p21 III- Expression des aquaporines en réponse à la lumière ... p21 DISCUSSION ET PERSPECTIVES ... p24 I- Méthodes et matériel végétal utilisé ... p24 II- Caractérisation de deux espèces de peuplier ... p25 1- Photosynthèse... p25 2- Conductance hydraulique foliaire ... p26 a- Réponse à la lumière... p26 b- Rythme endogène ... p26 c- Qualité de la lumière... p27 III- Etude de l’expression des PtPIP ... p28 1- En fonction des différents organes ... p28 2- En réponse à la lumière ... p29 Conclusion... p30 REFERENCES... p31 ANNEXE
Figure 1 : Schématisation des voies empruntées par l’eau dans les tissus végétaux
En rouge : la voie apoplasmique : l’eau reste dans l’espace extracellulaire.
En jaune : la voie symplasmique : l’eau emprunte les communications entre les cellules : les plasmosdesmes.
En bleu : la voie transcellulaire : l’eau passe par les membranes des différents compartiments qu’elle traverse.
Les fonctionnements hydraulique et photosynthétique des plantes ont été depuis de nombreuses années une source d’attention pour les chercheurs. La compréhension de ces phénomènes biologiques présente un grand intérêt fondamental avec de nombreux débouchés agronomiques. Les principales difficultés liées à ces études sont l’identification et la détermination de la contribution des différents acteurs entrant en jeu dans le fonctionnement et la régulation de ces phénomènes tout en sachant qu’il est difficile de les dé-corréler expérimentalement.
Les préoccupations actuelles concernent l’intégration des paramètres physiologiques et abiotiques avec la dynamique des conductances hydrauliques, l’établissement de modèles structures-fonctions, et l’influence du transport de l’eau sur les échanges gazeux.
I- Conductance hydraulique
L’eau est essentielle pour la plante qui absorbe beaucoup plus d’eau qu’un animal de poids identique (Raven, 2005). L’eau est un continuum dans la plante depuis les racines, où elle est prélevée du sol, jusqu’au limbe foliaire où elle est amenée via le xylème. Dans la feuille, l’eau va jusqu’aux stomates par où elle sort de la plante sous l’effet de la transpiration. Les mouvements d’eau au sein de la plante sont de trois types concourants. On distingue la voie apoplasmique, dans laquelle l’eau va se déplacer dans l’espace extracellulaire : les parois ; la voie symplasmique où l’eau va transiter en empruntant les voies de communications entre cellules végétales : les plasmodesmes. Enfin, l’eau peut circuler dans la plante en empruntant la voie transcellulaire et ainsi traverser les membranes (Figure 1). La conductance hydraulique est une grandeur mesurable qui est égale à l’inverse de la résistance exercée au passage de l’eau et correspond à la perméabilité d’un organe, d’un tissu ou même d’un type cellulaire vis-à-vis de l’eau. La conductance hydraulique de la plante peut se décomposer en trois éléments : la conductance racinaire (Kroot), la conductance de la tige (Kstem) et la conductance foliaire (Kleaf).
1- Conductance hydraulique et résistance foliaire
Kleaf est obtenue en faisant le rapport entre la transpiration (F) et une différence de potentiel hydrique entre la feuille (Ψleaf) et la tige (Ψstem). Rapportée à la surface de la feuille (S), Kleaf est exprimée en mmol d’H2O.m-2.s-1.Mpa-1.
Kleaf = F / ((Ψleaf –Ψstem)x S)
La résistance hydraulique foliaire (Rleaf) est égale à la somme des résistances de la voie xylémienne (Rxylem) et celles de la voie extra-xylémienne (Routside xylem). Ces deux résistances
A
B
Figure 2 : Fonctionnement de la photosynthèse (Campbell, 2005)
A- Principe schématique de la photosynthèse au sein de la cellule végétale
B- Principe schématique de la photosynthèse au sein des thylakoïdes du chloroplaste
Lors du transport d’électrons (flèches or), les protons du stroma traversent la membrane et s’accumulent dans l’espace intrathylakoïdien. Au moins trois des étapes des réactions photochimiques contribuent au gradient de protons : 1) le photosystème II scinde la molécule d’eau dans l’espace intrathylakoïdien ; 2) la plastoquinone (Pq) transfère les électrons au complexe de cytochromes de façon concomitante à une importation de protons dans l’espace intrathylakoïdien ; 3) le NADP+ capte deux protons dans le stroma lors de sa réduction en NADPH++H+. La
diffusion des protons de l’espace intrathylakoïdien vers le stroma alimente l’ATP synthétase. Déclenchées par la lumière, ces réactions emmagasinent l’énergie chimique dans le NADPH+H+ et dans l’ATP, qui apportent l’énergie
sont dynamiques. Deux hypothèses s’opposent pour expliquer l’origine de la résistance hydraulique foliaire totale et les contributions de Rxylem et Routside xylem : soit la résistance du xylème représente la majeure partie de la résistance foliaire, soit cette résistance du xylème est négligeable et la majeure partie de la résistance foliaire se trouve dans les tissus extra-xylémiens (Sack et Holbrook, 2006). Pour évaluer les contributions des résistances vasculaire et non-vasculaire, deux approches sont utilisées. La première consiste à procéder à un nombre important d’incisions au niveau des vaisseaux d’ordre supérieur à 3 pour éliminer la plus grande partie de la résistance non xylémienne (Sack et al., 2004). La deuxième méthode altère fortement et irrémédiablement la structure des membranes des cellules du mésophylle par traitement thermique : les feuilles sont bouillies ou congelées et décongelées (Tyree et al., 2001).
D’après les travaux de Cochard et al. (2004) effectués sur Laurus nobilis et Juglans
regia, les résultats des différents traitements visant à éliminer la contribution non xylémienne
convergent. Ces résultats indiquent une dominance de la résistance non xylémienne dans la résistance hydraulique totale. Ces résultats vont dans le sens d’une résistance vasculaire faible et d’une résistance mésophyllienne forte. Tyree et Cheung (1977) et Trifilò et al. (2003) ont montré, respectivement sur le hêtre et le tournesol, que la résistance extra-xylémienne représentait de 50 à 90 % de la résistance foliaire totale. Sack et al. (2004) ont montré la tendance inverse, à savoir que le xylème était responsable de 64 à 74 % de la résistance foliaire. Cette hypothèse est soutenue par les résultats obtenus sur le laurier par Zwieniecki et
al. (2002). Cette contradiction apparente dans les conclusions de ces travaux peuvent en partie
s’expliquer par des conditions expérimentales différentes.
2- Sources de variations de la conductance hydraulique foliaire
Cochard et al. (2000) ont montré que la résistance xylémienne changeait sous l’influence de la température à cause de la viscosité de l’eau. Celle-ci est également modulée par les modifications de teneur en ions de la sève xylémienne et augmentée avec le taux d’embolie. Les deux composantes de la résistance augmentent à faible température.
D’après Sack et al. (2005), les arbres des forêts tropicales ont leurs résistances xylémienne et extra-xylémienne qui sont corrélées linéairement entre les espèces. Lors de leurs expériences, Nardini et al. (2005) ont montré que la résistance extra-xylémienne est sous l’influence d’un rythme circadien chez le tournesol et qu’elle augmente à faible intensité lumineuse. Selon eux, la contribution de la résistance extra-xylémienne s’élève à près de 60 % de la résistance totale des feuilles au milieu de leur phase lumineuse et à 71,9 % pour les
Figure 3 : Cycle de Calvin (Hopkins, 2003)
Les enzymes représentées par des nombres inscrits dans des cercles sont : (1) la ribulose-1,5 bisphosphate carboxylase/oxygénase (Rubisco), (2) la 3-phosphoglycérate kinase et la glycéraldéhyde-3-phosphate déshydrogénase, (3) une aldolase, (4) la fructose-1,6 biphosphatase, (5) la transcétolase, (6) une aldolase, (7) la sédoheptulose-1,7- bisphosphatase, (8, 9) la ribulose-5-phosphate épimérase, (10) la ribose-ribulose-5-phosphate isomérase, (11) la ribulose-ribulose-5-phosphate kinase. Les chiffres entre parenthèse renseignent sur la stoechiométrie.
feuilles prises pendant la phase d’obscurité. Ces auteurs ont également montré que la restauration de Kleaf est plus rapide lors d’une inversion de photopériode dans le sens jour – nuit que lors de l’inversion réciproque (48 h contre 4 jours). Les feuilles de tournesol à l’obscurité ont une plus grande résistance extra-xylémienne que les feuilles à la lumière. Il est donc important de savoir comment la lumière induit une augmentation de la perméabilité à l’eau au niveau non vasculaire. Sôber a montré en 1997 qu’il existe chez Phaseolus Vulgaris une corrélation positive entre Kleaf et la conductance stomatique, et aussi que Kleaf était inférieure pour des plantes acclimatées à l’obscurité ou en manque d’eau par rapport aux plantes témoins.
Une hypothèse reste à vérifier : la variation de la conductance hydraulique foliaire pourrait être le résultat d’un processus actif qui aurait lieu dans les cellules vivantes du mésophylle, impliquant l’expression et ou l’activation d’aquaporines, protéines connues pour augmenter de façon significative la perméabilité à l’eau des membranes cellulaires (Eckert et
al., 1999).
II- Photosynthèse et conductance mésophyllienne au CO2 1- Photosynthèse
Le terme photosynthèse décrit deux phénomènes distincts : les réactions photochimiques et la fixation du CO2. Ces réactions ont lieu dans les tissus chlorophylliens de la plante : les feuilles essentiellement.
La phase photochimique a lieu pendant la journée. L’énergie lumineuse du soleil est captée par les pigments chlorophylliens insérés dans les membranes des thylakoïdes au niveau des chloroplastes. Cette énergie sert à générer un pouvoir réducteur et de l’ATP grâce à la photolyse de l’eau (Figure 2). Le pouvoir réducteur généré au cours de la journée sert à la fixation du CO2.
Le mécanisme au travers duquel le CO2 atmosphérique est incorporé aux glucides est le cycle photosynthétique de réduction du carbone (CPR) ou cycle de Calvin (Figure 3). On distingue chez les plantes plusieurs modalités de fixation du CO2. Les plantes comme le peuplier sont des plantes dites en C3 parce que le premier produit dans lequel le CO2 est incorporé par la Rubisco est un glucide à trois atomes de carbones : le 3-Phosphoglycérate (3-PGA). Chez ces plantes, un stress hydrique, même modéré va causer la fermeture des stomates et en conséquence, va limiter l’activité photosynthétique. Si la Rubisco est capable de fixer le CO2 au cours du cycle de Calvin, elle consomme de l’O2 et émet du CO2 aucours du cycle photosynthétique d’oxydation du carbone (CPO) ou photorespiration. Ce mécanisme
Figure 4 : Trajet du CO2 dans la feuille (Flexas et al., 2007)
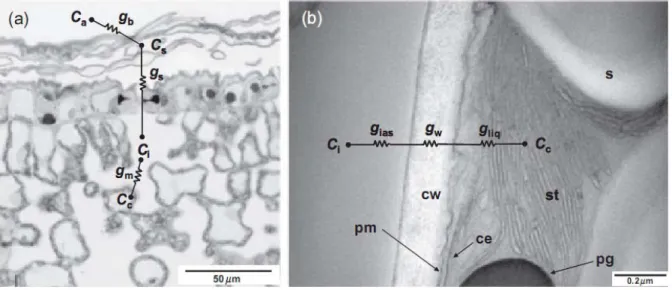
(a)- Observation en microscopie optique de la face inférieure de la feuille d’olivier. Le CO2 atmosphérique, à la concentration Ca rencontre une première résistance gb à l’approche de la surface de la feuille où sa concentration est définie par Cs. gb est connue sous le terme de couche limite. Les stomates exercent une résistance gs à l’entrée du CO2 dans la feuille. La concentration du CO2 dans la cavité stomatique ou l’espace intercellulaire est égale à Ci. La résistanceexercée au passage du CO2 entre l’espace intercellulaire et son site de fixation dans le stroma du chloroplaste est estimée par gm, la conductance mésophyllienne. La concentration en CO2 dans le stroma du chloroplaste est donnée par Cc
(b)- Observation en microscopie électronique d’une cellule du mésophylle d’une feuille de vigne. gm est décomposée en trois conductances : gias est la conductance de l’espaceintercellulaire, gw corresponds à la perméabilité de la paroi cellulaire, et gliq qui représente la conductance de la phase liquide de la cellule du cytoplasme au stroma du chloroplaste.
Pm : membrane plasmique, cw : paroi cellulaire, ce : enveloppe du chloroplaste, st : membrane des thylakoïdes, pg : plastoglobule, s : grain d’amidon.
serait une voie de dissipation du surplus énergétique reçu par les chloroplastes. L’assimilation photosynthétique nette (AN) est calculée comme la différence entre les produits de l’activité carboxylase et oxydase de la Rubisco et de la respiration mitochondriale.
AN = Vc – 0,5 V0 – Rd = Vc (1 - 0,5 V0/ Vc) - Rd
Vc : assimilation du CO par carboxylation du ribulose 1,5-bisphosphate
V0 : dégagement de CO2 par photorespiration
Rd : respiration mitochondriale
2- Conductance mésophyllienne au CO2
Le CO2 doit traverser plusieurs barrières physiques qui sont autant de résistances pour atteindre le lieu de sa fixation par la Rubisco (Figure 4). Le CO2 pénètre dans la feuille par l’ouverture créée quand les stomates sont ouverts : l’ostiole, ce qui signifie que la plante transpire. Le CO2 doit ensuite passer de l’espace intercellulaire à l’intérieur des cellules en franchissant la paroi et la membrane plasmique. Une fois dans le cytoplasme, le CO2 gagne le stroma du chloroplaste en franchissant la double membrane de l’organite. Le CO2 peut diffuser librement à travers les membranes biologiques, cependant pour assurer à la Rubisco, un approvisionnement suffisant en substrat, il est nécessaire d’envisager un mécanisme actif de transport du CO2.
Deux composantes vont limiter l’entrée du CO2. La première est la régulation de l’ouverture stomatique, donnée par la conductance stomatique (gs). La seconde est la conductance interne ou mésophyllienne (gm), qui renseigne sur la perméabilité au CO2 entre l’espace intercellulaire et le stroma. gm est obtenue en divisant l’assimilation nette par la différence de concentrations en CO2 de l’espace intercellulaire (Ci) et chloroplastique (Cc) :
gm = AN / (Ci -Cc)
La limitation de la photosynthèse par gm est souvent plus importante que celle par gs (Katsuhara et Hanba 2008). gm est corrélée positivement et linéairement aux rayonnement photosynthétiquement actifs (PAR ; Flexas et al., 2007b).
Terashima et Ono (2002) ont montré que gm diminuait en présence d’HgCl2. Les auteurs ont également montré un effet négatif de ce produit sur l’assimilation nette de CO2. Certaines aquaporines apparaissent comment des facteurs qui vont limiter ou faciliter le passage du CO2 à travers les membranes végétales et influencer la photosynthèse (Uehlein et al., 2003). Les aquaporines étant inhibées par le HgCl2, ces protéines semblent être de bonnes candidates pour expliquer gm. Ceci est corroboré par les travaux de Hanba et al. (2004) qui ont généré des lignées de riz surexprimant HvPIP2, une PIP2 d’Hordeum vulgare, et ont
Figure 5 : Structure schématique d’une aquaporine insérée dans une double membrane
(Hachez et al., 2006)
TM1 à 6 : domaines transmembranaires, HB et HE : hélices portant les domaines conservés NPA formant le pore. Les extrémités N et C terminales sont dans le cytoplasme.
constaté une augmentation de 40 % de gm par rapport au contrôle. Un effet positif sur gm a été observé chez le tabac par Flexas et al. (2006) en surexprimant l’aquaporine NtAQP1. Lorsque cette aquaporine était inhibée, un effet opposé a été observé par ces auteurs.
Les questions posées sont de savoir dans quelle mesure gm est influencée par l’activité d’aquaporines, d’identifier des isoformes impliquées et élucider des mécanismes de leurs régulations.
III- Les aquaporines
Découverte dans des érythrocytes humains par Peter Agre en 1992, la première aquaporine (AQP1) a été caractérisée comme un canal à eau par expression hétérologue dans des ovocytes de xénope. Les aquaporines appartiennent à la classe des Major Intrinsic Protein (MIP) et sont de petites protéines membranaires (300 acides aminés) présentes chez tous les organismes vivants des procaryotes aux animaux (Gomes et al., 2009).
Les MIP (Figure 5) sont caractérisées par la présence de six segments transmembranaires (TM), et deux motifs NPA (Asparagine, Proline et Alanine) et forment des pores dans les membranes. La structure primaire peut être divisée en deux moitiés similaires (TM1 à 3 et TM4 à 6) qui sont probablement apparues par duplication au cours de l’évolution (Zardoya, 2005). Au centre du pore, les domaines NPA forment de part et d’autre de la membrane de petites hélices hydrophobes se faisant face et participent à la sélectivité vis à vis des substrats (Hub et al., 2008). Le domaine NPA est chargé positivement ce qui provoque un réarrangement des molécules d’eau dans le pore. Au voisinage du domaine NPA se trouve la constriction aromatique/arginine (ar/R), qui est un filtre sélectif pour les substrats passant dans le pore (Beitz et al., 2006). Chez AQP1, 4 acides aminés forment la constriction ar/R dont l’histidine et l’arginine (H et R) qui agissent comme une interface hydrophile favorisant le transport en file simple des molécules d’eau (De Groot et Grubmuller, 2001). Les extrémités amine et carboxy-terminales sont du côté cytoplasmique et ont été identifiées comme importantes pour la régulation de l’ouverture du pore, des interactions protéines-protéines et de la perception osmotique (Chaumont et al., 2005).
Le génome humain compte 12 gènes codant des MIP, celui de la drosophile 7, et 11 chez Caenorhabditis elegans. Chez Arabidopsis thaliana, on dénombre 35 gènes contre 36 chez le maïs et 38 chez le riz. Leur nombre peut traduire l’importance des flux d’eau chez la plante comme l’absorption par les racines et la transpiration mais aussi le transport d’autres solutés.
Figure 6 : Distribution des aquaporines dans la cellule (Maurel et al., 2008)
La figure montre la localisation principale des MIP végétales caractérisées dans la littérature ainsi que les substrats qu’elles transportent et la direction des échanges (matérialisés par une flèche qui les traversent). Non présente sur cette illustration, NtAQP1, est une PIP1 du tabac localisée sur la membrane interne du chloroplaste avec une activité de transport du CO2 (Uehlein et al., 2008).
Il existe 4 familles de MIP chez les plantes (Kaldenhoff et al., 2008). Classées en fonction de l’homologie de leurs séquences avec celles qui ont été identifiées et caractérisées, leur nomenclature reflète leur localisation cellulaire (Figure 6). On distingue les Plasma membrane Intrinsic Protein (PIP), les Tonoplast membrane Intrinsic Protein (TIP), les Nodulin 26-like Intrinsic Protein (NIP) et les Small basic Intrinsic Protein (SIP).
1- Les différentes familles de MIP a- La famille des PIP
Caractérisées chez plusieurs espèces végétales comme des transporteurs d’eau, les PIP sont divisées en deux classes. Les PIP1 ont leurs extrémités N-terminales plus longues et leurs extrémités C-terminales plus courtes que celles des PIP2. Les expériences menées sur le maïs et le tabac suggèrent que les PIP1 ont une faible activité de transport de l’eau en comparaison des PIP2 quand elles sont exprimées seules dans les ovocytes de Xenopus laevis. Co-exprimée avec ZmPIP2 dans ce système, ZmPIP1;2 devient un transporteur fonctionnel d’eau et l’activité de transport est proportionnelle à la quantité de ZmPIP1;2 injectée (Fetter et al., 2004). Des résultats analogues ont été mis en évidence avec les MIP du tabac NtPIP1 ; 1 et NtPIP2;1 co-exprimées dans X. laevis (Mahdieh et al., 2008).
NtAQP1 est une PIP1 du tabac qui peut transporter le glycérol mais également le CO2. La modification de l’expression de cette dernière a un effet direct sur la conductance au CO2, la photosynthèse et la croissance de la plante (Uehlein et al., 2003 ; Flexas et al., 2006). La localisation sub-cellulaire de NtAQP1 par Uehlein et al. (2008) a permis de la mettre en évidence sur la membrane plasmique et sur la membrane interne du chloroplaste. Les PIP peuvent également transporter de l’H2O2 impliqué dans la signalisation cellulaire (Dynowski et al., 2008).
b- La famille des TIP
Les TIP sont essentiellement situées sur le tonoplaste, la membrane de la vacuole. La vacuole est une des particularités de la cellule végétale. Lieu de nombreuses réactions biochimiques, la vacuole est également le moteur de la croissance des cellules végétales en faisant pression sur la membrane plasmique. Les TIP sont les protéines les plus abondantes de cet organite (Maeshima, 2003). Elles ont été identifiées comme transporteurs d’urée, de glycérol, d’ammonium, d’H2O2 et de métaux lourds. Chez A. thaliana, la diminution de l’expression par ARN interférence de AtTIP1;1 provoque une sénescence accélérée, la mort
de la plante, une modification du métabolisme primaire et de la distribution d’assimilats alors que la teneur en eau de la plantule n’est pas altérée (Ma et al., 2004).
c- La famille des NIP
Les MIP de cette famille ont été nommées ainsi à cause de leur similarité avec la protéine nodulin-26 exprimée sur la membrane du symbiosome légumineuse-rhizobactérie. Elles sont subdivisées en 2 sous familles (NIP1 et NIP2), existent chez toutes les plantes et ont généralement une faible activité de transport de l’eau. Un rôle des NIP a été mis en évidence dans le transport de petits solutés comme le glycérol et l’urée. AtNIP5;1 exprimée chez le xénope révèle une activité de transporteur de bore. La carence en bore induit l’expression d’AtNIP5;1 et le mutant d’insertion nip5;1 montre un phénotype proche de celui causé par une carence en bore (Takano et al., 2006). AtNIP2;1 est un transporteur de l’acide lactique et est induit dans les racines en anoxie (Choi et Roberts, 2007).
d- La famille des SIP
Dans cette famille, les membres ont une faible similitude avec les autres MIP, environ 20 %. Ce sont également les moins nombreuses : 1 chez A. thaliana et le maïs et 2 chez la vigne. Les SIP sont également subdivisées en SIP1 et SIP2 et sont essentiellement associées aux membranes internes de la cellule notamment avec le réticulum endoplasmique (Ishikawa
et al., 2005). Peu d’informations sont disponibles sur les substrats des SIP mais l’hypothèse
actuelle suggère qu’elle agissent comme régulateurs de la taille et de l’osmolarité du réticulum endoplasmique (Gerbeau et al., 1999).
2- Expression des aquaporines
Chez les plantes, les aquaporines apparaissent comme des éléments centraux du transport de l’eau et du CO2. Les démonstrations des implications des aquaporines dans la physiologie de la plante sont de plus en plus nombreuses. Les membres de la famille des PIP sont considérés comme responsables de la plus grande partie de la conductance hydraulique au niveau cellulaire. En 2003, Aharon et al. montrent que la surexpression constitutive de l’aquaporine PIP1b d’Arabidopsis chez le tabac entraîne une augmentation du taux de croissance de la plante, de la transpiration, de la densité stomatale et de l’efficacité photosynthétique dans des conditions favorables de croissance. En revanche, la surexpression d’AtPIP1b n’a pas d’effet bénéfique pour des plantes qui sont soumises à un stress salin.
Chez l’orge, l’isoforme HvPIP2;1 est exprimée dix fois plus que les isoformes HvPIP1;3 au niveau des racines. Leurs expressions dans des ovocytes de xénope ont permis de montrer une augmentation marquée de l’activité de transport d’eau pour l’isoforme HvPIP2;1 uniquement (Katsuhara et Hanba, 2008). La surexpression constitutive de cette isoforme chez le riz augmente la conductance hydraulique racinaire, et la sensibilité au stress salin (Katsuhara et al., 2003). Fetter et al. (2004) nous montrent que ZmPIP1;2 ne peut être trouvée dans la membrane plasmique que si elle est co-exprimée avec une ZmPIP2 ce qui permet de supposer que l’hétéro-dimérisation est nécessaire pour l’adressage de la protéine du réticulum à son site d’action.
3- Les aquaporines : à l’interface entre le métabolisme carboné et le fonctionnement hydraulique ?
Chez les plantes, le rapport entre l’entrée de CO2 et la sortie de vapeur d’eau détermine l’efficience d’utilisation de l’eau (WUE). Cette grandeur agronomique correspond au rapport entre la biomasse produite et l’eau consommée et présente un intérêt majeur pour améliorer la résistance à la sécheresse des espèces cultivées (Condon et al., 2004). Pour optimiser l’entrée de CO2 pour la photosynthèse tout en minimisant leurs pertes en eau, les plantes doivent ajuster en permanence leur efficience hydraulique en modulant la résistance hydraulique de leurs différents organes.
Les aquaporines, de part leur implication dans le transport de l’eau et du CO2 chez les plantes pourraient expliquer une partie des variations de WUE, soit en faisant varier l’abondance de ces protéines, soit en changeant leur état d’activation afin d’augmenter leurs activités de transporteurs.
IV- Le peuplier, modèle d’étude
Appartenant à la famille des Salicaceae, le genre Populus compte 35 espèces. La culture du peuplier représente environ 240 000 ha en France ce qui la place au premier rang européen pour la surface cultivée (Conseil National du Peuplier). Utilisés principalement par les industries de l’emballage et de la papeterie, les peupliers le seront peut être davantage à l’avenir à cause de leur pouvoir de stockage du CO2 important, qui pourra être mis à profit dans des programmes de lutte contre les gaz a effet de serre. L’INRA développe des programmes d’amélioration des espèces sur le plan de la résistance aux pathogènes et aux insectes, de la productivité et de la production de biomasse à usages biotechnologiques : biocarburants, bioplastiques, médicaments (molecular pharming), etc…
En 2006, Populus trichocarpa est devenu le premier arbre dont le génome était entièrement séquencé. Choisi pour son petit génome (485 Mb), sa culture et sa multiplication aisées en font un modèle de choix pour les études menées sur les arbres. Le peuplier est également un végétal à croissance rapide, jusqu’à 5 m par an, ce qui permet d’observer plus facilement les effets des traitements influant sur la croissance et le développement. Enfin, la transformation génétique du peuplier est une technique maitrisée qui permet de tirer avantage de cette plante pour procéder à de la validation fonctionnelle de gènes candidats.
P. trichocarpa est une espèce qui a été largement caractérisée pour ses capacités
hydriques. En 1991, Bassman et Zwier ont montré des WUE différentes sur des populations réparties dans des zones à humidités différentielles. Les plus faibles valeurs étant observées pour des populations situées en zones humides. Les arbres peuplant ces stations humides ne présentaient pas de contrôle stomatique de la transpiration. Ces données sont en accord avec les travaux de Sparks et Black (1999) effectués sur 4 populations de P. trichocarpa. Ces auteurs ont par ailleurs associé la sensibilité à la sécheresse avec un contrôle stomatique de la transpiration limité.
V- Objectifs du stage
L’étude de ces phénomènes fondamentaux sur le peuplier, modèle ligneux, est indispensable à une bonne compréhension des mécanismes impliqués pour deux raisons principales :
- la mise en place d’une stratégie de régulation chez une plante pérenne est plus complexe que pour une plante non pérenne du fait de sa stabilité dans le temps. Une telle stratégie doit logiquement prendre en compte les variations biologiques rencontrées au cours de l’arrêt et de la reprise d’activité pendant l’hiver. Les travaux qui ont été effectués au cours de mon stage de Master 2ème année de Recherche au sein de l’équipe HYDRO de l’UMR PIAF (UMR 457, UBP-INRA), se basent pour une partie sur ceux obtenus sur Juglans regia par Cochard et ses collaborateurs (2007). Ces auteurs ont mis en évidence une corrélation entre l’augmentation de la conductance hydraulique foliaire à la lumière et l’augmentation des transcrits d’aquaporines JrPIP.
- le génome du noyer n’étant pas séquencé, le peuplier s’est imposé comme un bon candidat pour poursuivre et approfondir ces études d’écophysiologie couplées à la biologie moléculaire.
Les objectifs de mon stage étaient d’identifier les gènes d’aquaporines dans le génome de Populus trichocarpa, de préciser les organes dans lesquels des gènes étaient
préférentiellement exprimés, d’étudier les fonctionnements hydraulique et photosynthétique de deux espèces de peupliers (Populus trichocarpa et Populus nigra) dans le but de mettre en évidence une éventuelle régulation de la conductivité hydraulique foliaire en réponse à la lumière, et enfin d’étudier l’implication possible des aquaporines dans les variations de conductance hydraulique foliaire.
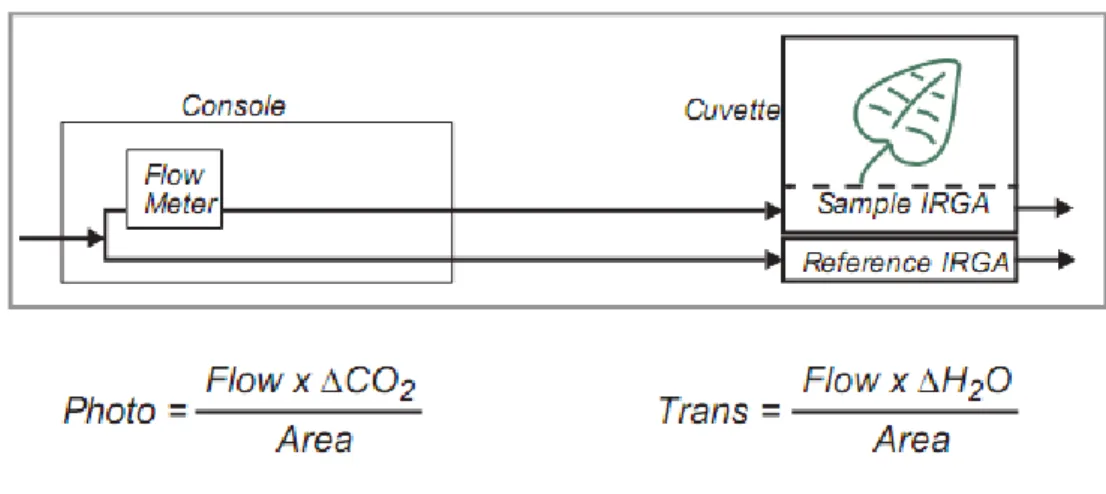
Figure 7 : Schéma du principe du LI-6400 (Using the LI-6400 v5.3 ,LI-COR)
La flèche représente le trajet de l’air qui parcoure le LI-6400. Une fois que la température de la feuille, le flux, le VPD, la concentration en CO2 dans l’échantillon, et l’irradiance sont réglées, les mesures effectuées par l’appareil sont :
Photo : Photosynthèse nette calculée à partir du flux d’air (Flow), et de la différence des
concentrations en CO2 entre l’air entrant et sortant de l’échantillon (∆CO2), autrement dit entre l’échantillon (Sample) et la référence. Cette différence est mesurée à l’aide des analyseurs de gaz à infrarouges (IRGA). Le résultat est ramené à la surface de l’échantillon contenu dans la chambre (Area, soit 6 cm2) et donné en moles de CO2.m-2.s-1.
Trans : Transpiration calculée à partir du flux d’air, de la différence des concentrations en eau
entre l’échantillon et la référence (∆H2O) et ramenée à une unité de surface. La transpiration est donnée en moles d’eau par m-2.s-1.
I- Matériel végétal
Deux espèces du genre Populus ont été étudiées : des clones de Populus nigra 71072-501 âgés d’un an ou de six semaines au moment des expériences et des clones de Populus
trichocarpa 101-74 âgés de six semaines. Le matériel végétal a été obtenu auprès de l’Unité
de Recherche Amélioration, Génétique et Physiologie Forestières (INRA d’Orléans). Les plantes ont été cultivées sous serre avec une humidité relative comprise entre 50 et 70 %, une température fixée à 22 °C le jour et 19 °C la nuit et un éclairage assuré par des lampes à sodium Master Son-T PIA 400W (Philips Electronics, N.V.). Le dispositif suppléait la lumière naturelle lorsque celle-ci était inférieure à 350 W.m-2 de 6 h à 23 h. L’irradiance à 40 cm mesurée à l’aide du LI-185B (LI-COR, Lincoln NE) est égale à 400 µmol.m-2.s-1.
II- Approches d’écophysiologie 1- Mesures de photosynthèse
a- Réponse au CO2
Lors des mesures, la température de la serre était fixée à 20 °C le jour et 18 °C la nuit, sous un éclairage continu de 6 h à 23 h par 4 lampes de 400 W Sylvania Britelux HSI/TSX (Havells Sylvania, Raunheim Germany) offrant 600 µmol.m-2.s-1 à 40 cm de la lampe. Les clones de P. nigra, d’environ 1 m étaient répartis en dessous sur une surface de 2 m2. Les clones P. trichocarpa mesurant 50 cm, ils ont été surélevés de 50 cm pour recevoir une irradiance égale. Cinq à 6 clones de chaque génotype ont été utilisés pour les expérimentations.
Les courbes de réponses au CO2 ont été obtenues grâce au système LI-6400 (LI-COR, Lincoln NE). Le LI-6400 calcule la photosynthèse en analysant par infrarouge la différence de composition entre les gaz contenus dans une chambre où est placé l’échantillon, et une référence (Figure 7). Les valeurs de photosynthèse nette sont obtenues à partir du modèle de Farquhar et al. (1980). Cet appareil permet de contrôler précisément l’ensemble des paramètres qui influent sur la photosynthèse dans une chambre de 6 cm2 où l’on place une partie de la feuille. Une source lumineuse de type LED rouge et bleue placée au dessus de l’échantillon permet de fixer l’irradiance à 1200 µmol.m-2.s-1, valeur non limitante pour le matériel végétal testé. Cette valeur, obtenue expérimentalement lors de travaux préalables de réponse à la lumière effectués sur le même matériel végétal, correspond au début de la saturation de la photosynthèse lors de l’application d’une irradiance croissante. La température de la feuille est fixée à 25 °C et le VPD (Vapor Pressure Deficit) est compris
Figure 8 : Schéma du principe du HPFM
De l’air exerce une pression de 2 Bars (0,2 MPa) sur une solution saline dont le trajet est symbolisé par les flèches et qui est perfusée dans deux feuilles testées en parallèle. Chaque feuille est reliée à un débitmètre (D1 et D2) qui mesure en permanence le flux de solution saline. Les feuilles sont placées dans un bain d’eau « pure » thermostaté à 25°C. Un ordinateur enregistre les valeurs des débitmètres et d’un manomètre afin d’obtenir les valeurs de conductance hydraulique foliaire. Le système permet de mesurer des changements de conductances hydrauliques foliaires au cours de différents traitements ou stimulations. Le schéma montre une stimulation lumineuse des feuilles. Il est possible d’intercaler entre la source lumineuse et les feuilles différentes sortes de filtres. Le système est également utilisable pour mesurer la réponse des feuilles à certains composés en leurs perfusant des solutions directement dans le circuit.
entre 1,2 et 1,6 kPa. Le LI-6400 permettant de contrôler la concentration en CO2 autour de l’échantillon, la concentration initiale est fixée à 350 µmol.mol-1 et la photosynthèse est mesurée. Des paliers sont successivement réalisés à 250, 150, 100, 50, et 600 µmol.mol-1. A partir de 600 µmol.mol-1, la concentration en CO2 a été augmentée par palier de 200 µmol.mol-1 jusqu’à observer une photosynthèse maximale.
b- Calculs des paramètres photosynthétiques
Le Vcmax (vitesse de carboxylation de la Rubisco)et le Jmax (vitesse de transport des électrons) sont estimés à partir d’ajustements des courbes d’assimilation ou photosynthèse (A) en fonction de la concentration en CO2 interne (Ci). Les données sont ajustées à l’aide du logiciel de statistiques R (voir Annexe) suivant les paramètres de Bernacchi et al. (2002) afin d’obtenir deux courbes de régression : la première s’alignant sur les points les plus proches de l’origine, sa pente indique le Vcmax à 25 °C ; la seconde s’alignant sur le plateau de points, sa pente indique le Jmax à 25 °C.
2- Mesure de la conductance hydraulique foliaire
L’appareil utilisé pour réaliser ces mesures est du type HPFM (High Pressure Flow Meter) et a été développé par l’INRA (Figure 8). Il s’agit d’un réservoir de solution saline sous une pression de 0,2 MPa (2 Bars) qui envoie la solution sous pression dans le pétiole de deux feuilles prélevées extemporanément. Les flux de solution saline sont mesurés entre le réservoir et les pétioles par un débitmètre Type E (Brokhorst High Tech, Ruurlo NL). La conductance hydraulique foliaire est calculée en faisant le rapport entre le flux d’eau qui passe par la feuille et par la pression appliquée pour faire passer cette eau. La solution passant à travers la feuille est dégazée afin de diminuer le plus possible la quantité d’air dissous et contient 1 mM de CaCl2 et 10 mM de KCl afin de s’approcher au maximum de la salinité de la sève brute. La feuille à étudier était placée dans un bain d’eau thermostaté à 25°C à moins d’un cm de la surface. La pression et les flux sont enregistrés par ordinateur toutes les 5 secondes.
a– Etude de variations en réponse à la lumière
Placée à 40 cm de la surface de l’eau, une lampe de 400 W Sylvania Britelux HSI/TSX éclaire les feuilles. L’irradiance dans le laboratoire est proche de 0 et est égale à 600 µmol.m-2.s-1 au niveau de la feuille lorsque la lampe est allumée. La différence de réponse
Tableau I : Amorces utilisées pour le clonage des aquaporines
Nom Séquence UTR 5’
Tm (°C) Séquence UTR 3‘ Tm (°C) Taille ADNc (pb) PtPIP1-A AGATCTGATATCCTTTCRCTGTTARCGCTGCA 60 CGTATTGCTATTATTACTTGTGT 48 1067
PtPIP1-B AGATCTGATATCCAAACMANCAACCCATCTTAC 59 TTCAAAAAGCTAGATAATTTACAC 47 1035
PtPIP1-C AGATCTGATATCTAATTGTTWATWCTTAATACTGCG 57 ACGTCACTGTTATAGCACGC 52 1057
PtPIP1-D AGATCTGATATCCAAATACACTTAGCTTAAGCTATAA 59 GAGTAATTTACATAAATGGAACC 48 1155
PtPIP1-E AGATCTGATATCTGACTGAGAAAGGCCAGAGGA 63 AAGAATTCACTGAGAGGGAATA 49 1129
PtPIP2-A AGATCTGATATCACACAGTTACAAGCCGAGAG 62 GTGACGGAGTGAGTAAAAGAGTG 55 1010
PtPIP2-B AGATCTGATATCCATTTCTCCACACTTCCACC 62 AGCTCACAACCATGACATAAC 50 1079
PtPIP2-C AGATCTGATATCCTTTTCTTTGTAAGAAGAACCTA 59 ACCGAAAGGGATAATAAAGGG 50 1023
PtPIP2-D AGATCTGATATCCTCTTCTTCTGTCAACCCTTC 62 CCACCCCACGAGAGAAATC 53 1030
PtPIP2-E AGATCTGATATCCTTTAGCTACCATAACACTTAC 60 ACACATTCCCCAGTTCCAAG 59.8 1047
PtPIP2-F AGATCTGATATCCTACATTTGAAGTTGAGGTAGC 61 ACATAACATGACCCAACTGAG 50 1063
PtPIP2-G AGATCTGATATCCGTCGCAAACCAGTAGCTAG 63 CCGTGACAATGATACACAGCA 52 1093
PtPIP2-H AGATCTGATATCTGCGCTCTAAGAACCACAGAG 63 ACCCACATATCAAGTTGAGAC 50 1026
PtPIP2-I AGATCTGATATCGGAGAAGAAAGATAGTTTGGC 61 GATTCACTTTGCCCACCTTG 52 1134
T7 TAATACGACTCACTATAGGG 48
entre une feuille prise sur une plante non acclimatée à l’obscurité et pour une autre prise sur une plante acclimatée à l’obscurité est mesurée.
b- Recherche d'une régulation endogène de la conductance hydraulique
Afin de mettre en évidence un rythme endogène de régulation de la conductance hydraulique, certaines plantes ont été acclimatées à l’obscurité complète pendant 48 h. Des prélèvements ont été effectués au cours de la journée sur P. nigra (à 8 h, 10 h, 12 h, 14 h et 18 h). Pour chaque point on compare la réponse à la lumière d’une feuille prise sur une plante contrôle non acclimatée à l’obscurité et d’une autre prise sur une plante acclimatée à l’obscurité. Les feuilles ont été choisies pour avoir des surfaces foliaires les plus proches possibles pour chaque prélèvement et chaque échantillon a été prélevé sur le 4ème rang foliaire en partant de l’apex.
c- Etude du rôle de la qualité de la lumière
La lumière est filtrée par un film anti-bleu doublé obtenu auprès de Didier Combes de l’Unité d'Ecophysiologie des Plantes Fourragères (INRA de Lusignan). Le filtre utilisé avec la lampe Sylvania de 400 W laisse passer un rayonnement lumineux de 200 µmol.m-2.s-1 contre 600 µmol.m-2.s-1 sans le filtre à 40 cm de la lampe. La diminution de l’irradiance au niveau de la feuille mesurée sous le filtre rouge est répercutée à la feuille témoin pour laquelle un filtre neutre est appliqué. Ce filtre neutre diminue avec la même intensité toutes les longueurs d’ondes, contrairement à l’anti bleu qui diminue plus fortement les longueurs d’ondes correspondant au bleu que les autres.
III- Approches de biologie moléculaire 1- Séquences des aquaporines
Les séquences d’aquaporines putatives ont été obtenues lors d’une recherche exhaustive dans le génome de Populus trichocarpa disponible sur le portail du DOE Joint Genome Institute (www.jgi.doe.gov). Les séquences ont ensuite été confrontées aux séquences d’EST du genre Populus (NCBI : www.ncbi.nlm.nih.gov/BLAST/).
2- Clonage des aquaporines
A partir d’un pool d’ADNc issus de différents organes de Populus trichocarpa disponible au laboratoire, et en utilisant des amorces situées dans les UTR 5’ et 3’ (Tableau I), les séquences des aquaporines de la famille des PIP ont été insérées dans le vecteur
p-GEM®T-Figure 9 : Cartes des vecteurs utilisés
A- Carte du vecteur pGEM®-T Easy (www.promega.com/vectors/t_vectors.htm).
B- Carte du vecteur d’expression T7TS (www.xenbase.org/xenbase/original/www/Marker_pages
/PlasMaps/T7TS.eps)
Tableau II : Amorces utilisées pour l’amplification par qPCR des aquaporines
Nom Séquence 5’ Tm (°C) Séquence 3’ Tm (°C) Taille ADNc (pb) Taille ADNg (pb) PtPIP1-A AAGAAGTGATTATTATGATTATGG 47 GATTTGAAGAAACACGTATTGCTA 51 168 168
PtPIP1-B TTCAAGAGCAGAGCTTAATTTC 49 TTCAAAAAGCTAGATAATTTACAC 47 154 154
PtPIP1-C CAAGAAGTGATGAYTGTGATGC 50 ACGTCACTGTTATAGCACGC 52 144 144
PtPIP1-D GACAGAAAACAAGGTTCTTTAGA 50 GAGTAATTTACATAAATGGAACC 48 178 178
PtPIP1-E TCGTTCCCAATCRATGGATGTT 50 AAGAATTCACTGAGAGGGAATA 49 155 155
PtPIP2-A 2 TGGTACACTTGGCCACAATC 59.4 GCTAATGCTCCCACAAATGG 60.5 142 258
PtPIP2-B ATTCAGTTATGGAACGTACGC 50 AGCTCACAACCATGACATAAC 50 118 118
PtPIP2-C CCAACGGTTTTAAATCTCGGTTC 53 ACCGAAAGGGATAATAAAGGG 50 105 105
PtPIP2-D 2 ATCCCAATCACAGGAACTGG 59.8 CTATGGCTGCACCAATGAAG 59.3 127 223
PtPIP2-E ATATGGTCACTGAAATACTTCC 49 CCCAATACGCGTACAAACGT 52 130 130
PtPIP2-F 2 ATGACCATTGGCTCTTCTGG 60.1 GGCTTTAAGCATTGCTCCTG 60 126 126
PtPIP2-G TGCTAACATGGTGGCTCCC 53 GATCATCCCAGGCTTTCTTATC 53 272 798
PtPIP2-H GCAATCCTACTAGCTAAGGC 52 ACCCACATATCAAGTTGAGAC 50 155 155
PtPIP2-I 2 ACTGGAACTGGCATCAATCC 59.9 CCAACCCAGAAAATCCACAG 60.3 98 381
18S GCGATTTGTCTGGTTAATTCCG 53 GGCCAAGGCTATAGACTCG 53 209
EF1α GACAACTAGGTACTACTGCACTGTC 58 TTGGTGGACCTCTCGATCATG 54 236 B
Easy (Promega, Madison WI) et clonées dans des bactéries compétentes JM109 (Réf. L1001 Promega, Madison WI). Une fois clonées dans p-GEM®T-Easy, les séquences ont été amplifiées avec l’amorce universelle T7 et une amorce anti-sens spécifique de l’isoforme clonée pour vérifier la nature, la taille de l’insert ainsi que son insertion dans la bonne orientation. Les séquences des amorces 5’ commencent par AGATCT suivit de GATATC qui sont respectivement les sites de restriction des enzymes Bgl II et Eco RV, l’ajout de ces séquences permet ainsi de digérer les inserts afin de les sous-cloner dans le vecteur T7TS (A. Johnson et P. Krieg) (Figure 9). Les clones ont été vérifiés par séquençage SP6 et T7 (Cogenics, Meylan France).
3- PCR quantitative en temps réel a- Amorces utilisées
Les amorces de PCR quantitative ont été dessinées à l’aide du logiciel Primer3Plus, (Untergasser et al., 2007 ; www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi). Les amorces de PCR quantitative utilisées (Tableau II) doivent amplifier des séquences de 100 à 200 pb, situées le plus proche possible de l’extrémité 3’UTR et idéalement toutes avoir une Tm voisine de 59 °C. Les températures de fusion réelles des amorces correspondent aux Tm calculées sur le site de Promega (www.promega.com/biomath/calc11.htm).
Les amorces ont ensuite été confrontées par BLAST aux banques d’EST du genre
Populus disponibles sur le site du NCBI pour ne conserver que des amorces spécifiques d’une
isoforme mais aspécifiques dans le genre Populus ceci afin de pouvoir amplifier les isoformes chez les différentes espèces de ce genre.
Les amorces ont également été validées en PCR classique en utilisant comme matrice de l’ADN génomique de P. trichocarpa et puis avec des ADNc issus de P. trichocarpa, P.
nigra et P. tremula alba. L’efficacité et la spécificité des amorces ont été vérifiées d’une part
au cours de PCR en gradient de température de 10 °C autour du Tm théorique et d’autre part avec des dilutions d’ADNc de P. trichocarpa.
b– Extraction d’ARN et reverse transcription
Les ARN totaux des différents échantillons ont été extraits suivant le protocole décrit par Chang et al. (1993). La quantité et la qualité des ARN totaux a été évaluée par dosage spectrophotométrique à 260 nm à l’aide du Nanodrop ND-1000 (NanoDrop products, Wilmington, DE, USA) puis par dépôt d’1 µg d’ARN totaux sur gel d’agarose à 1,5 %. La
synthèse d’ADNc simple brin à été faite à partir d’1 µg d’ARN totaux avec le kit SuperScript III First-Strand Synthesis System (Invitrogen, Carlsbad CA).
c- Matériel utilisé et programmes PCR
Les PCR en temps réel ont été réalisées à l’aide du thermocycleur iCycler iQ (Bio-Rad, Hercules CA) dans un volume réactionnel de 30 µl en utilisant 2 µl d’ADNc dilués au vingtième et 0,1 µl de Platinum Taq Polymerase (Invitrogen, Carlsbad CA).
Deux programmes ont été utilisés suivant la sous-famille des PIP (PIP1 ou 2) à laquelle appartiennent les transcrits dont l’expression était suivie. Les réactions commençaient par 3 minutes à 94 °C pour la dénaturation initiale suivies de 40 cycles de : 15 secondes de dénaturation à 94 °C, 15 secondes d’hybridation à 54 °C pour les PIP1 et 57 °C pour les PIP2 et 20 secondes d’extension à 72 °C. La réaction était terminée par une dissociation des produits de PCR de 57 ou 54 °C jusqu’à 94 °C par pas de 0,5 °C pour s’assurer que les échantillons n’étaient pas contaminés et que la réaction s’était bien déroulée.
d- Profils d’expression tissus spécifique des PIP1 et 2 chez P. trichocarpa
Les ADNc de bourgeons, de l’écorce et du bois, de feuilles à différents stades de développement et des racines d’un clone de P. trichocarpa ont été amplifiés par PCR en utilisant les amorces de PIP1 et PIP2. Les threshold cycle (Ct) bruts ont tout d’abord été standardisés en utilisant l’ARN ribosomique 18S comme contrôle interne et les valeurs obtenues ont été exprimées en unités arbitraires à partir de l’équation :
Valeur = - 0,0615 x Ct + 2,25
Cette formule correspond à l’équation d’une droite étalon qui associe au Ct le plus précoce (cycle 20) la valeur « 1 » et au Ct le plus tardif (cycle 40) la valeur « 0 ». Les résultats présentés correspondent à une seule répétition biologique avec 3 répétitions techniques (deux répétitions pour chaque gène à l’intérieur de chaque répétition technique).
e- Profils d’expression en réponse à la lumière des PIP1 et 2 chez P. trichocarpa et P. nigra
Pour évaluer les différences d’expression des gènes d’aquaporines en réponse à la lumière, les trois premières feuilles complètement développées ont été prélevées sur P. nigra et P. trichocarpa. Les 8 plantes utilisées avaient été préalablement placées à l’obscurité pendant 48 h. Le prélèvement témoin correspond au retour à la lumière. Différents prélèvements ont été réalisés après 15 et 30 minutes ,1 et 2 heures sous éclairement de 400
µmol.m-2.s-1. Les plantes ont été ensuite replacées à l’obscurité et des prélèvements ont été réalisés au bout de 1, 2 et 4 h d’obscurité. Les feuilles ont été immédiatement congelées dans de l’azote liquide puis les ARN ont été extraits puis reverse transcrits. Deux microgrammes d’ARN totaux ont servi de matrice pour la reverse transcription. Les variations d’expression ont été mises en évidence par la méthode du ∆∆Ct
4- Arbre phylogénique
Un arbre phylogénétique a été généré grâce au site Phylogeny (www.phylogeny.fr) à partir des séquences protéiques prédites des MIP putatives de Populus trichocarpa et les 35 MIP d’Arabidopsis thaliana en utilisant les paramètres par défaut du logiciel (Dereeper et al., 2008).
Figure 10 : Arbre phylogénique réalisé à partir des séquences protéiques prédites des 63 MIP putatives de P. trichocarpa et des 35 MIP d’A. thaliana
Réalisé à partir sur le site phylogeny.fr avec les valeurs par défaut. Les valeurs en rouge sur les branches correspondent à la probabilité que les séquences se trouvent à cet endroit dans l’arbre.
I- Caractérisation des aquaporines de Populus trichocarpa
L’étude des banques de données a permis d’identifier 63 MIP putatives chez P.
trichocarpa. L’analyse à l’aide d’un algorithme de phylogénie moléculaire des séquences
protéiques prédites a permis de mettre en évidence que 10 de ces MIP sont homologues avec les PIP2 d’Arabidopsis thaliana, 6 peuvent être rattachées aux PIP1, 20 aux TIP, 13 aux NIP, 7 aux SIP, et 7 séquences se distinguent remarquablement des familles de MIP décrites jusqu’à présent (Figure 10). Ce nombre fait de Populus le genre ayant le plus de gènes codant pour des MIP. En terme de répartition, 25 % de ces gènes codent des PIP contre 36 % chez
Arabidopsis, environ 30 % codent des TIP et 20 % codent des NIP chez les deux genres. Les
SIP sont assez bien représentées chez Populus qui en compte 3,5 fois plus qu’Arabidopsis. Le peuplier est donc un modèle d’étude qui apparait comme plus complexe. Il a été choisi du fait de la disponibilité de l’intégralité de son génome dans les bases de données. Ceci pour avoir une vision globale de l’expression des gènes codant des MIP au cours des traitements considérés et pouvoir les associer à des réponses biologiques. Pour limiter l’étendue du champ d’investigation, nous avons restreint notre étude aux PIP1 et 2 qui sont décrites comme transporteurs d’eau et de CO2 dans la littérature. Les premiers travaux qui ont été réalisés sont des profils d’expression tissus-spécifiques de 14 des PIP putatives sur P. trichocarpa par qPCR.
1- PIP1
Cette étude nous renseigne sur le lieu préférentiel d’expression des différentes isoformes. On peut ainsi constater que la plupart des PIP qui sont suivies sont plutôt ubiquitaires dans les organes étudiés.
On retrouve systématiquement dans le pool des PIP1 dominantes, l’isoforme PtPIP1-A et PtPIP1-C (Figures 11 A et B). Celles-ci sont exprimées avec une intensité comparable dans les différents organes. L’isoforme PtPIP1-E, qui est moins exprimée est également assez stable dans les tissus étudiés à l’exception de la feuille sénescente où son expression n’est pas détectée. L’isoforme PtPIP1-B, dont l’expression globale est assez stable, semble cependant être légèrement plus exprimée dans la tige avec un maximum observé pour le xylème. L’isoforme PtPIP1-D apparaît comme n’étant pas du tout exprimée au niveau des feuilles. Cette dernière est également absente dans la tige en formation, elle est faiblement exprimée dans les racines et apparaît comme plus spécifique de la tige en croissance et surtout des bourgeons.
Figure 11 : Niveaux d’expression des PtPIP1 chez un P. trichocarpa âgé de 6 semaines par qPCR
Les amplifications ont été réalisées à partir d’ADNc de différents organes de P. trichocarpa dilué au 1/20è en utilisant les amorces du tableau II avec le programme décrit dans la section « matériels et méthodes ».
A- Contribution exprimée en pourcentage de l’expression des différentes PtPIP1 en fonction des organes.
B- Expression relative des PtPIP1 en fonction des organes en unités arbitraires, l’écart type correspond à 4 répétitions des réactions de PCR.
Figure 12. Niveaux d’expression des PtPIP2 chez un P. trichocarpa âgé de 6 semaines par qPCR
Les amplifications ont été réalisées à partir d’ADNc de différents organes de P. trichocarpa dilué au 1/20è en utilisant les amorces du tableau II avec le programme décrit dans la section « matériels et méthodes ».
A- Contribution exprimée en pourcentage de l’expression des différentes PtPIP2 en fonction des organes.
B- Expression relative des PtPIP2 en fonction des organes en unités arbitraires, l’écart-type correspond à 4 répétitions des réactions de PCR.
2- PIP2
PtPIP2-A, PtPIP2-C et PtPIP2-G sont les isoformes les plus fortement exprimées quelques soient les organes considérés et à des niveaux relativement comparables (Figures 12 A et B). PtPIP2-A a une expression plus importante dans la tige où sa contribution notamment dans le xylème où elle représente plus de 20 % de l’expression des PIP2. PtPIP2-D est particulièrement stable entre tous les organes observés. Parmi les 9 PIP2 dont l’expression a été suivie, certaines montrent des profils d’expression plus contrastés. L’isoforme PtPIP2-I est très peu exprimée dans les feuilles et la tige en développement mais relativement abondante dans les bourgeons, la tige en croissance et surtout les racines. Au niveau de la tige, on observe une augmentation des transcrits entre le stade juvénile et le stade adulte où l’on observe des compositions en PIP2 très proches entre xylème et phloème.
3- Au niveau des feuilles
Il est intéressant de remarquer qu’au cours de la croissance des feuilles, on observe une augmentation de l’expression relative de la plupart des transcrits de PIP1 et 2 suivie d’une diminution globale au cours du vieillissement. Ainsi, PtPIP1-A ; B ; E ; et F et PtPIP2-B ; D ; E ; F suivent ce profil d’expression.
Exception faite de PtPIP2-C et I qui sont assez stables au cours de l’ontogenèse de la feuille, l’expression des PIP2 est dynamique. Au cours de la croissance et du développement de la feuille, le pool de transcrits semble augmenter parallèlement avec l’augmentation des activités biologiques.
D’après ces résultats, l’isoforme PtPIP1-D n’est pas exprimée dans les feuilles et PtPIP2-I l’est très peu. Ce genre d’information est important dans la mesure où les travaux qui sont mis en œuvre ont pour objectif de corréler des activités biologiques qui ont lieu dans les feuilles avec l’expression des MIP. L’ensemble des PIP de P. trichocarpa étudiées ont été clonées dans le vecteur p-GEM®T-Easy afin de vérifier par séquençage la spécificité des amplifications mais également en prévision de tests de fonctionnalité dans le système Xenopus laevis. Les gènes candidats à ces tests seront dans un premier temps sous clonés dans le vecteur d’expression T7TS.
II- Caractérisation de deux espèces du peuplier : Populus trichocarpa et Populus
nigra
Afin de caractériser les deux espèces de peuplier, des expériences de photosynthèse ont été réalisées.
A B
Figure 13 : Courbes de réponse au CO2 de P. nigra et P. trichocarpa
Courbes représentatives AN = f (Ci) obtenues après traitement des enregistrements du LI-6400 par l’algorithme d’ajustement des courbes qui détermine Jmax et Vcmax en utilisant les paramètres de Bernacchi et al. (2002).
Tableau III : Valeurs moyennes de Jmax et Vcmax de P. nigra et P. trichocarpa.
Vcmax Jmax
(µmol CO2.m -2
.s-1) Ecart-type (µmol d’électrons.m-2.s-1) Ecart -type
P. nigra 55,01 3,5 112,42 1,98
P. trichocarpa 41,53 5,53 104,41 2,67
1- Photosynthèse
Les facteurs qui influencent la photosynthèse sont nombreux. La culture en serre permet de contrôler au mieux les conditions de culture et de limiter les variations des facteurs influant sur la photosynthèse et leur impact. La température est fixée, un éclairement constant et une bonne alimentation en eau est offert aux plantes qui sont espacées afin de ne pas se créer mutuellement de l’ombre. Les deux genres de Populus qui servent de modèles d’études dans le travail ci-présent ont des efficacités photosynthétiques différentes. La caractérisation des facteurs internes à la plante, influençant la photosynthèse est préalablement nécessaire à la modélisation de la photosynthèse en impliquant des MIP pour la conductance mésophyllienne au CO2. Le LI-6400 est un système d’analyse de la photosynthèse qui fait référence auprès des chercheurs et des professionnels de l’agronomie. Il permet notamment de fixer la concentration en CO2 à des valeurs inférieures ou supérieures à celle de l’atmosphère. Chacun des deux genres a été testé 5 à 6 fois en utilisant des plantes différentes pour s’affranchir de la variabilité biologique. Celle-ci est supposée faible dans la mesure où il s’agit de clones de même âge, cultivés dans les mêmes conditions. Ces mesures ont été réalisées à différentes heures de la journée sans différence significative ce qui écarte une influence quantifiable d’un rythme biologique endogène sur la conductance mésophyllienne au CO2.
Les courbes AN= f(Ci) nous indiquent que lorsque l’irradiance n’est pas limitante pour la feuille, alors on peut observer une saturation de la photosynthèse pour une concentration interne en CO2 (Ci) de 600 µmol.mol-1 (Figures 13 A et B). Le point d’intersection de la courbe avec l’axe Ci est le point de compensation pour le CO2 à 1200 µmol.m-2.s-1 et correspond à la concentration en CO2 pour laquelle l’assimilation est égale à sa production par photorespiration et respiration mitochondriale. Cette valeur se situe aux alentours de 50 µmol.mol-1 pour les deux espèces étudiées.
Les valeurs de Jmax et Vcmax est données par l’algorithme d’ajustement à la courbe avec les valeurs de Bernacchi et al. (2002). Les résultats présentés dans le Tableau III montrent que les paramètres estimés avec l’hypothèse gm infinie sont identiques pour les deux espèces de peupliers testées.
Flexas et al. (2007b) ont mis en évidence une variation importante de gm au cours du vieillissement de la plante. Ces auteurs montrent en effet une diminution de 28 % de gm ayant pour conséquence une diminution de 41 % d’AN. Il se pourrait qu’une partie de la variation observée au cours des différentes mesures trouve son origine dans de petites variations de l’âge de la feuille malgré les précautions prises pour un échantillonnage homogène.
Figure 14 : Evolution de Kleaf en fonction du temps et
de la lumière chez Populus
trichocarpa mesurée par
HPFM sous 2 bars de pression
A - Effet du rang foliaire. Le rang n+1 (rouge) montre une augmentation de Kleaf de 55 %
contre 30 % pour la feuille du rang supérieur.
B - Effet de l’acclimatation d’une plante à l’obscurité pendant 24 h par rapport au témoin. La feuille acclimatée (rouge), présente une augmentation de 32 % de Kleaf
contre 0 % pour la feuille non acclimatée (verte) de même rang. C - Effet de l’acclimatation d’une plante à l’obscurité pendant 72 h par rapport au témoin. Plus d’augmentation de Kleaf n’est
observable. Le Kleaf de la feuille
acclimatée (rouge) diminue au cours du temps.
D - Effet de la composition du
rayonnement lumineux. L’utilisation d’un filtre anti bleu
entre la feuille et la source lumineuse de 12 à 13 h induit une augmentation de Kleaf de 35 %
contre 19 % pour la feuille masquée par le filtre opaque noir. En abscisse : période d’obscurité (bleu), période de transition (noir), période de lumière (jaune)