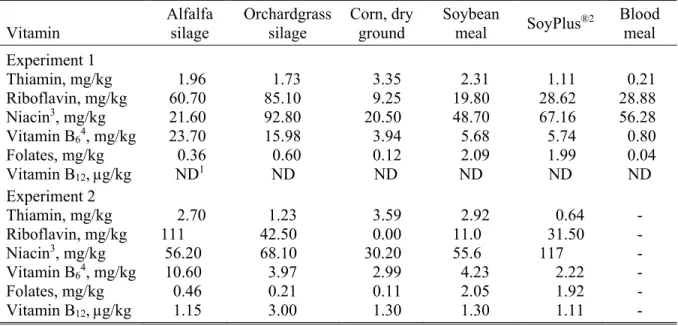
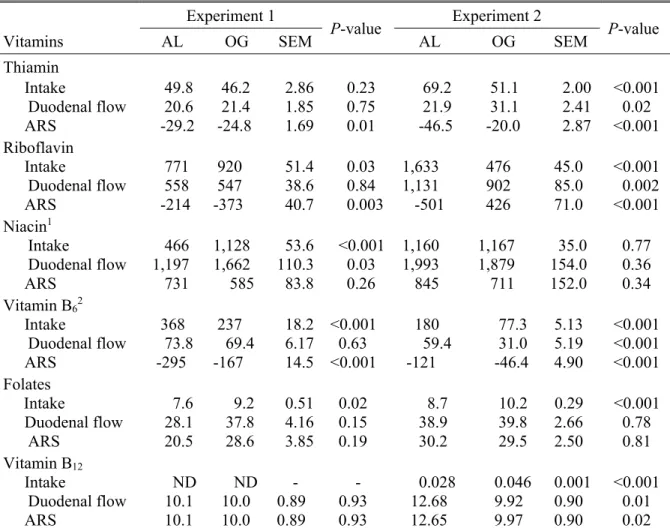
Vitamines B, éléments clés de l'efficacité
métabolique : Effets de la nature de la diète sur les
apports
Thèse
Douglas de Souza Castagnino
Doctorat en sciences animales
Philosophiae doctor (Ph.D.)
Résumé
Les travaux sur la nutrition en vitamines B des ruminants montrent des résultats très variés sur les quantités de ces nutriments disponibles pour l’animal selon la nature de la ration. Ces divergences sont dues à des changements des populations microbiennes dans le rumen, causées par les facteurs physico-chimiques de la ration. Une amélioration de la compréhension des effets de la nature de la diète sur la synthèse et l’utilisation des vitamines B dans le rumen pourrait aider à identifier les conditions sous lesquelles une supplémentation en ces vitamines serait bénéfique pour la vache. Le but de ce travail de thèse est donc d’améliorer la compréhension des effets de l’espèce fourragère, de la maturité et de la longueur des particules de fourrage sur les apports en vitamines B chez la vache laitière. Pour évaluer chacune de ces variables, les concentrations de thiamine, riboflavine, niacine, vitamine B6, folates et vitamine B12 ont été mesurées dans les échantillons d’aliments et de digesta duodénal recueillis lors de trois projets réalisés à l’Université du Michigan par l’équipe du Dr. M. Allen. Dans la première étude, l’effet de l’espèce fourragère des ensilages a été évalué au cours de deux expériences similaires durant lesquelles les vaches recevaient une diète à base d’ensilage de luzerne ou de dactyle. Les diètes à base de luzerne ont été associées à une augmentation de la dégradation de la thiamine et de la vitamine B6 dans le rumen par rapport aux diètes à base d’ensilage de dactyle. La deuxième étude visait à évaluer les effets de la maturité des plantes lors de la mise en silo sur les quantités de vitamines B disponibles pour la vache; les deux expériences se différenciaient par l’espèce fourragère étudiée, soit la luzerne ou le dactyle. Une récolte à un stade de maturité plus élevé a augmenté les flux duodénaux de thiamine, de niacine et de folates lorsque les vaches recevaient des diètes à base d’ensilage de luzerne mais n’a diminué que le flux duodénal de riboflavine chez les animaux recevant des diètes à base d’ensilage de dactyle. La troisième étude a comparé les effets de la longueur de coupe (10 vs. 19 mm) d’ensilages de luzerne et de dactyle sur le devenir des vitamines B dans le système digestif de la vache laitière. Cette étude a permis de constater qu’une augmentation du temps de séchage au champ diminuait les concentrations de vitamines B dans les ensilages. Cependant, la taille des particules des ensilages de luzerne et de dactyle n’a pas affecté les quantités des vitamines B arrivant au duodénum des vaches. En général,
les résultats de ces études montrent qu’il existe une corrélation négative entre la synthèse de riboflavine, de niacine et de vitamine B6 et leur ingestion, suggérant une possible régulation de la quantité de ces vitamines B par les microorganismes du rumen. De plus, l’ingestion d’amidon et d’azote a été corrélée positivement avec la synthèse de thiamine, de folates et de vitamine B12, et négativement avec la synthèse de niacine. Ces corrélations suggèrent que les microorganismes qui utilisent préférentiellement l’amidon jouent un rôle majeur pour la synthèse ou la dégradation de ces vitamines. De plus, la présence d’une quantité suffisante d’azote semble avoir un impact majeur sur ces processus. La suite de ces travaux devrait viser la modélisation de ces données afin de mieux appréhender la physiologie de la digestion de ces vitamines et permettre la création de modèles mathématiques capables de prédire les quantités de vitamines disponibles pour les vaches. Ces modèles permettront, lorsqu’intégrés aux logiciels de formulation de ration, d’élaborer une diète plus précise, ce qui améliorera la santé du troupeau et la performance laitière et augmentera les profits du producteur.
Abstract
Research on B vitamins and ruminant nutrition reported that the amounts of these nutrients available for the animals vary according to the nature of the diet. These differences are due to changes in microbial populations in the rumen caused by several physical and chemical characteristics of the diets. However, data regarding how those factors affect B-vitamin supply are limited. A better quantitative understanding of dietary factors driving ruminal synthesis and degradation of B vitamins will help to identify conditions favoring vitamin supplementation to the cow. The objective of this thesis was to evaluate the effects of forage family, plant maturity and forage particle length on B-vitamin supply to lactating dairy cows. To evaluate each of these variables, concentrations of thiamin, riboflavin, niacin, vitamin B6, folates and vitamin B12 were measured in feed and duodenal digesta samples collected during three studies carried out at the University of Michigan by Dr. Allen’s team. In the first study, the effect of forage family was evaluated in two similar trials, where diets containing either alfalfa or orchardgrass silages were offered to lactating dairy cows. Diets based on alfalfa silage resulted in a greater degradation of thiamin and vitamin B6 in the rumen compared with diets based on orchardgrass silages. The second study assessed the effects of plant maturity at harvest on the amounts of B vitamins available to the cow; the two trials differed by the studied forage family, alfalfa or orchardgrass. Increasing maturity at harvest of forages increased the amounts of thiamin, niacin and folates reaching the sites of absorption with diets based on alfalfa silages whereas it only decreased riboflavin duodenal flow with diets based on orchardgrass silages. In the third study, the effects of alfalfa and orchardgrass silage particle length (10 vs. 19 mm) on the fate of B vitamins in digestive tract of dairy cows were studied. Increasing wilting time in the field decreased B-vitamin concentrations in both silages. Nevertheless, for both forage families, the length of silage particles had no effect on the amounts of B vitamins reaching the duodenum of cows. Overall, the results from these three studies highlighted that there was a negative correlation between the apparent synthesis of riboflavin, niacin and vitamin B6 and their intake, suggesting that ruminal bacteria reduced the synthesis of these vitamins when their dietary supply was augmented. In addition, starch and nitrogen intakes were positively correlated with the
ruminal synthesis of thiamin, folates and vitamin B12, but negatively correlated with the synthesis of niacin. These correlations suggest that microorganisms that use starch play a major role in the synthesis or the degradation of these vitamins. Moreover, these processes are affected by the nitrogen supply. The continuation of this work will be the modeling of these data, which will help to better understand the digestion physiology of these vitamins and allow the creation of mathematical models able to predict duodenal B-vitamin flow in cows. These models will allow feed formulation software to design more adequate diets, which will improve herd health and milk performance and increase dairy farmer profits.
Table des matières
Résumé……….. ... iii
Abstract……… ... v
Table des matières ... vii
Liste des figures ... xiii
Liste des tableaux ... xv
Liste des abréviations ... xvii
Remerciements ... xxi
Avant-propos ... xxiii
1. INTRODUCTION ... 1
2. REVUE DE LITTÉRATURE ... 5
Structure chimique et rôles métaboliques ... 8
2.1.1. Thiamine (vitamine B1) ... 8 2.1.2. Riboflavine (vitamine B2) ... 9 2.1.3. Niacine (vitamine B3) ... 10 2.1.4. Vitamine B6 ... 12 2.1.5. Folates (vitamine B9) ... 13 2.1.6. Vitamine B12 ... 14 Synthèse ruminale ... 16 2.2.1. Rapport fourrage:concentré ... 16 2.2.2. Niveau d’ingestion ... 17
2.2.3. Taille des particules ... 17
2.2.4. Source de glucides et niveau d’azote ... 17
2.2.5. Les additifs et le cobalt ... 18
Analyse des vitamines B ... 21
2.4.1. Chromatographie en phase liquide à haute performance (CLHP) ... 22
2.4.2. Méthode microbiologique pour la détermination des folates ... 24
2.4.3. Méthode de radio-dosage pour la détermination de vitamine B12... 25
3. ARTICLE 1 : ESPÈCE FOURRAGÈRE / FORAGE FAMILY ... 27
Résumé ... 31
Abstract ... 33
Introduction ... 35
Materials and methods ... 36
3.4.1. Experiment 1 ... 36
3.4.1.1. Cows and treatments ... 36
3.4.1.2. Sample and data collection ... 38
3.4.2. Experiment 2 ... 38
3.4.2.1. Cows and treatments ... 38
3.4.2.2. Sample and data collection. ... 38
3.4.3. Laboratory analyses ... 38 3.4.3.1. Thiamin ... 39 3.4.3.2. Riboflavin ... 40 3.4.3.3. Niacin ... 41 3.4.3.4. Vitamin B6 ... 42 3.4.3.5. Folates ... 43 3.4.3.6. Vitamin B12 ... 43
3.4.4. Calculations and statistical analyses ... 44
3.5.2. Apparent ruminal synthesis and duodenal flow ... 46 Discussion ... 49 3.6.1. Thiamin ... 49 3.6.2. Riboflavin ... 50 3.6.3. Niacin ... 51 3.6.4. Vitamin B6 ... 51 3.6.5. Folates ... 52 3.6.6. Vitamin B12 ... 52 Conclusions ... 53 Acknowledgments ... 53 References ... 54
4. ARTICLE 2 : MATURITÉ DE LA PLANTE / PLANT MATURITY ... 59
Résumé ... 63
Abstract ... 65
Introduction ... 67
Material and methods ... 68
4.4.1. Treatments and cows ... 68
4.4.1.1. Trial 1 ... 70
4.4.1.2. Trial 2 ... 70
4.4.2. Sample and data collection ... 70
4.4.3. Laboratory analyses ... 70
4.4.4. Calculations and statistical analyses ... 71
Results ... 72
4.5.1. Intake, apparent ruminal synthesis and duodenal flow ... 73
Conclusions ... 80
Acknowledgments ... 80
References ... 81
5. ARTICLE 3 : LONGUEUR DES PARTICULES / PARTICLE LENGTH ………...……….85
Résumé ... 89
Abstract ... 91
Introduction ... 93
Materials and methods ... 94
5.4.1. Treatments and cows ... 94
5.4.1.1. Trial 1 ... 96
5.4.1.2. Trial 2 ... 96
5.4.2. Sample and data collection ... 96
5.4.3. Laboratory analyses ... 96
5.4.4. Calculations and statistical analyses ... 97
Results and Discussion ... 99
5.5.1. Daily intake, apparent ruminal synthesis and duodenal flow ... 99
Conclusions ... 106
Acknowledgements ... 106
References ... 107
6. DISCUSSION GÉNÉRALE / CONCLUSION ... 109
Synthèse apparente de vitamines B dans le rumen et paramètres alimentaires et ruminaux ... 113
6.1.1. Influence du type de microorganismes sur le bilan de vitamines dans le rumen……… 114
6.1.2. Influence du taux de croissance des microorganismes et influence du
substrat sur le bilan vitaminique du rumen ... 116
Besoins en vitamines B chez les vaches laitières... 118
Limites de l’étude ... 119
Conclusion ... 120
Liste des figures
Figure 2.1 : Fonctions métaboliques des vitamines B (adaptée de Le Grusse et Watier, 1993). ... 7
Figure 2.2 : Structure chimique de la thiamine et de ses dérivés phosphorylés. ... 9
Figure 2.3 : Structure chimique de la riboflavine et de ses dérivés. ... 10
Figure 2.4 : Structure chimique de la niacine et de ses dérivés. ... 11
Figure 2.5 : Structure chimique de la pyridoxine et de ses dérivés. ... 12
Figure 2.6 : Structure chimique de l’acide folique et de ses dérivés. ... 14
Figure 2.7 : Structure chimique de la cobalamine et de ses dérivés. ... 15
Figure 2.8. Représentation schématique de l’analyse des vitamines. ... 21
Figure 2.9. Exemple théorique d’un chromatogramme produit par une méthode d’analyse de vitamines par CLHP en phase inverse. ... 23
Liste des tableaux
Tableau 2.1. Reproductibilité inter- et intra-essai et pourcentage de recouvrement ... 23 Table 3.1. Ingredients, nutrient composition and B-vitamin concentrations of the two treatment diets containing alfalfa silage (AL) or orchardgrass silage (OG) in experiments 1 and 21 ... 37
Table 3.2. B-vitamin concentrations (mg/kg DM) of ingredients used in the experimental diets ... 44 Table 3.3. Intake, duodenal flow and apparent ruminal synthesis (ARS) of B vitamins (mg/d) in responses to diets based on alfalfa (AL) or orchardgrass (OG) silages (LS Means) in experiments 1 and 2
... 45
Table 3.4. Duodenal flow and apparent ruminal synthesis (ARS) of B vitamins (mg/kg DMI) in responses to diets based on alfalfa (AL) or orchardgrass (OG) silages (LSMeans) in experiments 1 and 2 47
Table 3.5. Pearson correlations coefficients between apparent ruminal synthesis (ARS) of B vitamins and dietary and digestive parameters ... 48
Table 4.1. Ingredients, chemical composition and B-vitamin concentrations of treatment diets containing alfalfa or orchardgrass harvested and ensiled as early- or late-cut1 ... 69
Table 4.2. B-vitamin concentrations (mg/kg DM) of ingredients used in the experimental diets containing alfalfa or orchardgrass harvested and ensiled as early- or late-cut ... 72
Table 4.3. Intake, duodenal flow and apparent ruminal synthesis (ARS) of B vitamins in response to diets containing alfalfa or orchardgrass harvested and ensiled as early- or late-cut ... 74
Table 4.4. Duodenal flow and apparent ruminal synthesis (ARS) of B vitamins in response to diets containing alfalfa or orchardgrass harvested and ensiled as early- or late-cut ... 75
Table 4.5. Pearson correlations coefficients between apparent ruminal synthesis (ARS, mg/d) and dietary and digestive parameters combining data from both experiments ... 77
Table 5.1. Ingredients, nutrient composition and B-vitamin concentrations of the two treatment diets containing alfalfa or orchardgrass both chopped to two different particle lengths1 ... 95
Table 5.2. B-vitamin concentrations (mg/kg DM) of ingredients used in the experimental diets containing alfalfa or orchardgrass silages both chopped to long or short particles1 ... 98
Table 5.4. Duodenal flow and apparent ruminal synthesis (ARS) of B vitamins (mg/kg of DMI) in response to diets containing alfalfa or orchardgrass silages both chopped to long or short particles ... 101
Table 5.5. Pearson correlations coefficients between apparent ruminal synthesis (mg/d) and dietary and digestive parameters combining data from both experiments ... 103
Tableau 6.1. Coefficients de corrélation de Pearson entre la synthèse apparente des vitamines B dans le rumen et les paramètres alimentaires et digestifs combinant les données des trois expériences ... 114
Liste des abréviations
AL = alfalfa
ADN = acide désoxyribonucléique ARS = apparent ruminal synthesis CC = coupe courte
CH = coupe hâtive CL = coupe longue
CLHP = chromatographie en phase liquide à haute performance Co = cobalt CP = crude protein CT = coupe tardive d = day DA = dactyle DF = duodenal flow DM = dry matter
DMI = dry matter intake EC = early-cut
FAD = flavine adénine dinucléotide FD = Flux duodénal
FMN = flavine mononucléotide h = hour (s)
HPLC = high performance liquid chromatography iNDF = indigestible neutral detergent fiber
j = jour
LC = late-cut (dans le chapitre 4) LC = long-cut (dans le chapitre 5) LS means = least square means LU = luzerne
MO = matière organique MS = matière sèche
MSI = matière sèche ingéré N = nitrogen
NA = nicotinic acid
NAD = nicotinamide adénine dinucléotide
NADP = nicotinamide adénine dinucléotide phosphate NAM = nicotinamide
ND = not-detected
NDF = neutral detergent fiber NS = not-significant min = minute (s) OG = orchardgrass OM = organic matter P5P = pyridoxal-5’-phosphate PAL = pyridoxal PAM = pyridoxamine PL = particle length PLP = phosphate de pyridoxal PYR = pyridoxine
SAR = synthèse apparente dans le rumen SC = short-cut
SE = standard error
SEM = standard error of the means TMP = thiamine monophosphate TMR = total mixed ration
TPP = thiamine pyrophosphate TTP = thiamine triphosphate VFA = volatile fatty acid
« Science works on the frontier between knowledge and ignorance. We’re not afraid to admit what we don’t know or we were wrong about this. There’s no shame in that. The only shame is to pretend that we have all the answers. »
Remerciements
J’aimerais adresser ma profonde gratitude à Christiane Girard, ma co-directrice. Son expérience scientifique et sa générosité m'ont permis de développer considérablement mes aptitudes de chercheur et de comprendre les enjeux du domaine. Mes plus sincères remerciements vont également à Rachel Gervais, ma directrice. Rachel, vous avez une capacité exceptionnelle à captiver les personnes avec votre spontanéité et votre énergie contagieuse. À vous deux, merci aussi d’avoir rendu possible mon stage à l’Université de Cornell.
I am grateful to Thomas Overton and the members of his research group for hosting me during my research internship at Cornell University. It was an enriching experience both from the scientific and personal point of view.
I want to thank Dr. Michael Allen for his collaboration in all three studies, providing the samples for the B vitamin analysis and contributing to article publications.
J’exprime ma gratitude envers le Programme de recherche en partenariat pour l’innovation en production et transformation laitière Novalait - Agriculture et Agroalimentaire Canada - Fonds de recherche du Québec Nature et Technologies - Ministère de l’Agriculture, des Pêcheries et de l’Alimentation du Québec, Québec, Canada pour m’avoir attribué ma bourse d’étude et subventionné mes études au doctorat.
Je tiens également à remercier chaleureusement Débora Santschi, Yvan Chouinard et Jonh Metcalf pour avoir accepté d’être examinateurs externes dans mon comité de thèse.
Chrystiane Plante, merci pour votre aide technique essentielle au laboratoire, pour votre ouverture d'esprit et vos bons conseils. Merci aussi au Centre de Recherche et de Développement de Sherbrooke - Agriculture et Agroalimentaire Canada de m’avoir accueilli et fourni l’infrastructure nécessaire à la conduite des analyses laboratoires.
Un merci spécial à Valérie Beaudet (100% québécoise) de me faire découvrir la culture et une bonne partie des beaux paysages de sa province. Les allers à l’Université Laval et les retours à Sherbrooke ont été mémorables.
Merci à mes nombreux collègues, autres étudiants, qui m’ont aidé à parcourir ce long chemin qu’est la thèse.
« Si j'ai vu plus loin, c'est en montant sur les épaules de géants », Isaac Newton. Ces
géants sont mes parents, Círia et Angelo Castagnino, et mon frère, Pablo Castagnino, qui m’ont donné tout le support nécessaire pour que je puisse réaliser mes études supérieures loin du « chez nous ». À vous, mes plus sincères gratitudes.
Plus personnellement, je remercie la personne qui me fait rêver… Sophie, qui me fait regarder le monde avec d’autres yeux. Merci pour cet accent, pour être toujours là quand j’ai besoin et pour partager ta vie avec moi.
Avant-propos
Ce travail de thèse vise à étudier l’impact des caractéristiques de la diète sur la synthèse apparente des vitamines B dans le rumen de la vache laitière. Cette thèse comprend une mise en contexte (Chapitre 1), une revue des travaux antérieurs (Chapitre 2), trois articles scientifiques (Chapitres 3 à 5), et une discussion/conclusion (Chapitre 6). L’auteur de cette thèse est l’auteur principal de chacun des articles. Sa participation aux projets consistait à analyser les concentrations de thiamine, riboflavine, niacine, vitamine B6, folates et vitamine B12 dans des échantillons d’aliments et de digesta duodénaux récoltés lors de projets réalisés par M. Michael Allen (Michigan State University), ainsi qu’à analyser les données, rédiger les articles et participer au processus de soumission et de révision des articles, sous la supervision de ses codirecteurs de thèse, Mme Rachel Gervais et Mme Christiane Girard.
Chapitre 3 : D. S. Castagnino, M. Seck, V. Beaudet, K. L. Kammes, J. A. Voelker Linton, M. S. Allen, R. Gervais, P. Y. Chouinard and C. L. Girard. 2016. Effects of forage family on apparent ruminal synthesis of B vitamins in lactating dairy cows. Journal of Dairy Science 99:1884-1894. doi: 10.3168/jds.2015-10319.
Chapitre 4 : D. S. Castagnino, K. L. Kammes, M. S. Allen, R. Gervais, P. Y. Chouinard and C. L. Girard. Plant maturity affects apparent ruminal synthesis of B vitamins in lactating dairy cows. Animal, soumis.
Chapitre 5 : D. S. Castagnino, K. L. Kammes, M. S. Allen, R. Gervais, P. Y. Chouinard and C. L. Girard. Particle length of silages affects apparent ruminal synthesis of B vitamins in lactating dairy cows. Journal of Dairy Science, publié le 19 avril 2016.
À la fin des années 20, Bechdel et ses collaborateurs (1926; 1928) fournissaient des preuves concluantes quant à la capacité de la microflore du rumen à produire des vitamines B en quantités suffisantes pour permettre aux bovins de se développer et de produire une descendance même en présence d’une alimentation apportant peu de ces nutriments. En 1954, Kon et Porter (1954) ont démontré que les concentrations de vitamines B dans le rumen de l’animal étaient relativement constantes. À l’époque, les quantités de ces vitamines issues des régimes alimentaires et de la synthèse microbienne dans le tractus digestif des ruminants étaient suffisantes pour ne pas provoquer de symptômes de carences chez des animaux possédant un rumen fonctionnel et bien développé. L’ensemble de ces informations a contribué à diffuser un dogme stipulant que les ruminants n’ont pas besoin de supplémentation en vitamines B. Cependant, depuis quelques années, le concept de «nutriment limitant», mis de l’avant par les travaux sur les besoins en acides aminés, a gagné en popularité. Ce concept a amené une remise en question sur les besoins de la vache laitière en vitamines B, dans le but d’optimiser l’efficacité de leur métabolisme ainsi que leurs performances zootechniques (Girard, 2008).
Au cours des trois dernières décennies, plusieurs études ont démontré que les vaches laitières hautes productrices pouvaient bénéficier de suppléments de vitamines B (thiamine: Shaver et Bal, 2000; niacine: Fronk et Schultz, 1979, Riddell et al., 1981; biotine: Majee et al., 2003, Zimmerly et Weiss, 2001; acide folique et vitamine B12: Girard et Matte, 1998, Preynat et al., 2009). Ces articles suggèrent une amélioration de l’efficacité métabolique quand il y a une supplémentation de ces vitamines. En production laitière, cette amélioration induit, d’une part, un meilleur bilan environnemental et, d’autre part, une diminution des coûts de production. D’un autre côté, le plus grand inconvénient de cette supplémentation est sa forte dégradation dans le rumen pouvant varier de 68% à 99% (Santschi et al., 2005a). De plus, probablement liées à cette dégradation variable dans le rumen, il n’y a pas toujours de réponses aux suppléments de vitamines B (Rosendo et al., 2004; Girard et al., 2005) et les résultats observés pour une vitamine ne peuvent pas être extrapolés aux autres vitamines B.
Chez les ruminants, la performance de l’animal dépend aussi bien de l’apport de vitamines par le régime alimentaire que de la synthèse de vitamines dans le rumen. La
synthèse peut être affectée par divers facteurs alimentaires et, par conséquent, changer les quantités de vitamines disponibles pour l’animal. Parmi les facteurs réputés affecter la physiologie de la digestion, les principaux sont le niveau d’ingestion totale (Sutton et Elliot, 1972; Zinn et al., 1987), le rapport fourrage:concentré (Voelker Linton et Allen, 2008), le type et le niveau de maturité du fourrage (Kammes et Allen, 2012a, b), la nature et le traitement des ingrédients (Miller et al., 1989a), la taille des particules (Kammes et Allen, 2012c) et les additifs (Miller et al., 1989b). Ces facteurs peuvent aussi bien modifier le pH ruminal, le temps de mastication et de rumination, que le taux de dégradation et le temps de transit et, par conséquent, ils peuvent altérer la fermentation. Pourtant, malgré ces connaissances, la quantité d’information disponible dans la littérature scientifique concernant les effets de ces facteurs sur les quantités de vitamines B disponibles pour l’animal est très limitée.
L’hypothèse de ce projet de recherche était qu’il est possible d’identifier les facteurs nutritionnels permettant de prédire les quantités de vitamines B disponibles pour la vache laitière. Le premier objectif de cette thèse était d’évaluer les apports alimentaires, la synthèse apparente dans le rumen et le flux duodénal de thiamine, de riboflavine, de niacine, de vitamine B6, de folates et de vitamine B12 selon la nature de la diète. Le second objectif était d’identifier les facteurs alimentaires ayant le plus d’influence sur les quantités de vitamines B disponibles pour la vache. Les trois principaux facteurs alimentaires évalués étaient : (1) l’espèce fourragère, comparant la luzerne vs le dactyle; (2) la maturité de la plante, en comparant une coupe hâtive vs une coupe tardive et; (3) la longueur de coupe des ensilages, en comparant une coupe de 10 mm vs 19 mm de longueur.
2. REVUE DE
LITTÉRATURE
Les vitamines sont un groupe de composés organiques présents en quantités minimales dans les aliments. Elles ont un faible poids moléculaire et n’ont pas de valeur énergétique. Cependant, elles jouent un rôle essentiel dans le fonctionnement normal de l’organisme, et des apports insuffisants peuvent provoquer des symptômes de carence (ex., selon les vitamines, signes généraux, neuropsychiques, cardiaques, cutanéomuqueux, oculaires, etc.). Les vitamines ne peuvent pas être synthétisées par les animaux (à l’exception de la niacine, et des vitamines C et D, selon les conditions et l’espèce animale).
Les vitamines du complexe B sont hydrosolubles et sont regroupées pour des raisons historiques, celles-ci ayant été initialement isolées dans les mêmes sources, généralement le foie et/ou les levures. Chaque vitamine présente une structure chimique et des rôles métaboliques différents. Les principales fonctions des vitamines B sont d’agir comme cofacteur pour les enzymes du métabolisme des acides aminés, des acides gras, de l’énergie et des acides nucléiques et d’être impliquées dans des réactions d’oxydo-réduction et dans la transcription de gènes (Figure 2.1).
Les huit vitamines B reconnues sont la thiamine (B1), la riboflavine (B2), la niacine (B3), l’acide pantothénique (B5), le complexe B6 (pyridoxal, pyridoxamine, pyridoxine), la biotine (B8), les folates (B9) et la cobalamine (B12). Le devenir des vitamines B5 et B8 ne sera pas traité dans cette thèse.
Structure chimique et rôles métaboliques
2.1.1. Thiamine (vitamine B1)
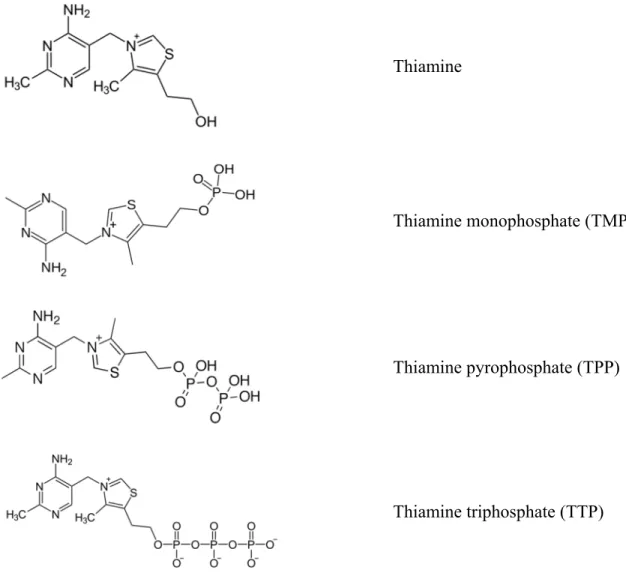
Connu pour sa carence causant le béribéri chez l’homme, une maladie qui affecte les nerfs et le cœur, le 3-[(4-amino-2-methyl-5-pyrimidinyl)methyl]-5-(2-hydroxyethyl)-4-methylthi-azolium (IUPAC), aussi appelé la vitamine B1, aneurine ou simplement thiamine, fut le premier facteur nutritionnel responsable de maladies carentielles à être isolé, identifié puis synthétisé en 1911 par C. Funk. Cette vitamine est une molécule composée d’un cycle pyrimidique et d’un cycle thiazolique soufré et azoté (Le Grusse et Watier, 1993). Il existe trois dérivés phosphorylés de la thiamine : mono, di, et triphosphates (Figure 2.2).
La thiamine pyrophosphate est un cofacteur essentiel pour l’organisme vivant et joue un rôle clé dans le métabolisme des acides aminés et des glucides (Begley et Ealick, 2010). Elle agit comme un coenzyme pour trois décarboxylases des acides α-cétoniques et pour deux transcétolases.
Contrairement aux plantes et aux microorganismes, les animaux sont incapables de synthétiser la thiamine et doivent donc compter sur des sources alimentaires exogènes. Chez les bovins, des carences de cette vitamine, notamment dues à la présence dans le rumen de l’enzyme thiaminase bactérienne (par exemple, Clostridium sporogenes et quelques espèces de Bacillus), qui détruit ou altère la thiamine, peuvent causer la poliencéphalomacie (i.e. nécrose du cortex cérébral). Les aliments pour animaux riches en thiamine sont le tourteau de soja, l’orge, le son de blé, la levure de brasserie déshydratée, le son de riz et le maïs (Schwab et al., 2006; Sauvant et al., 2004).
Thiamine
Thiamine monophosphate (TMP)
Thiamine pyrophosphate (TPP)
Thiamine triphosphate (TTP)
Figure 2.2 : Structure chimique de la thiamine et de ses dérivés phosphorylés.
2.1.2. Riboflavine (vitamine B2)
La riboflavine est le nom trivial donné au composé 7,8-dimethyl-10-(1’-D-ribityl) isoalloxazine, anciennement connu sous le nom de vitamine B2, vitamine G ou lactoflavine. Initialement confondue avec les autres vitamines du groupe B, la riboflavine fut la deuxième vitamine du complexe B découverte. La riboflavine résulte de la combinaison d’une flavine qui est un hétérocycle azoté à 3 noyaux et d’un sucre à 5 carbones, le ribose (Le Grusse et Watier, 1993). Les formes métaboliquement actives sont généralement appelées flavine mononucléotide (FMN) et flavine adénine dinucléotide (FAD; Figure 2.3).
Riboflavine Flavine mononucléotide (FMN)
Flavine adénine dinucléotide (FAD)
Figure 2.3 : Structure chimique de la riboflavine et de ses dérivés.
La riboflavine est essentielle pour le métabolisme intermédiaire des glucides, des acides aminés et des lipides. Plus précisément, les coenzymes FMN et FAD sont impliquées lors de réactions d'oxydo-réduction des acides gras et dans des réactions du cycle de Krebs. D’une part, la FMN est liée à l’activité des enzymes L-aminoacide oxydase, lactate déshydrogénase et 5’-phosphate oxydase pour l’activation de la vitamine B6. D’autre part, la FAD est liée aux enzymes D-aminoacide et glucose oxydases et succinyl et acétyl-CoA déshydrogénases, à la coenzyme cytochrome réductase et à la glutathion réductase.
Les principales sources alimentaires pour les ruminants présentant des fortes concentrations en riboflavine sont le foin de graminées, le foin de luzerne, la farine de poisson et la levure de brasserie déshydratée (Sauvant et al., 2004; Schwab et al., 2006).
2.1.3. Niacine (vitamine B3)
La niacine, nom générique de l’acide pyridine 3-carboxylique, aussi connue comme la vitamine PP ou vitamine B3, est le nom donné à deux molécules qui ont toutes deux une activité vitaminique, l’acide nicotinique et le nicotinamide. Cette dernière est le précurseur de deux dérivés importants sur le plan métabolique : (1) la nicotinamide adénine
la présence d’un radical phosphorylé sur le carbone 2’ du ribose associé à l’adénine (Le Grusse et Watier, 1993; Figure 2.4).
La niacine fut isolée en 1867, mais son importance ne fut établie que 70 ans plus tard, quand son rôle sur la prévention de la pellagre (i.e. maladie chez l’homme se manifestant par des dermatites, des diarrhées et de la démence) fut établi. En 1945, il fut démontré chez le rat que le tryptophane pouvait prévenir la carence de cette vitamine. Quelques années plus tard, la synthèse d’acide nicotinique à partir du tryptophane fut démontrée chez cette même espèce (Le Grusse et Watier, 1993).
La niacine participe à plusieurs réactions métaboliques sous la forme de nicotinamide, NAD et NADP. Ces deux dernières sont utilisées par plus de 30 déshydrogénases aussi bien dans le métabolisme des glucides (ex. : glucose-6-phosphate déshydrogénase), des lipides (ex. : α-glycérol-phosphate déshydrogénase) et des protéines (ex. : glutamate déshydrogénase) que dans le cycle de Krebs et la synthèse de la rhodopsine (avec l’alcool déshydrogénase; Combs Jr., 2012).
Acide nicotinique Nicotinamide Nicotinamide adénine dinucléotide (NAD) Nicotinamide adénine dinucléotide phosphate (NADP)
Les principales sources alimentaires de niacine sont, pour les ruminants, les pâturages, les coques de soja, la farine de poisson, la levure de brasserie, le tourteau de tournesol et le tourteau de canola (Sauvant et al., 2004). Cependant, la niacine présente dans les céréales est généralement peu disponible pour l’organisme animal (FAO, 1993).
2.1.4. Vitamine B6
La vitamine B6 est le nom générique de tous les dérivés du 3-hydroxy-2-méthylpyridine ayant une activité biologique chez le rat (Combs Jr., 2012). Ce groupement peut être une fonction alcool (pyridoxol ou pyridoxine), aldéhyde (pyridoxal) ou amine (pyridoxamine; Le Grusse et Watier, 1993; Figure 2.5).
Pyridoxine Pyridoxal Pyridoxamine Phosphate de pyridoxal
Figure 2.5 : Structure chimique de la pyridoxine et de ses dérivés.
Les trois formes, pyridoxine, pyridoxal et pyridoxamine, peuvent être phosphorylées. Par exemple, la forme phosphorylée du pyridoxal est le phosphate de pyridoxal (PLP). Celui-ci a un rôle métabolique particulièrement important car il est la forme qui sert de coenzyme à plus de 140 enzymes dont la plupart sont impliquées (1) dans le métabolisme des acides aminés (ex. amine transférases, désaminases, décarboxylases, désulfhydrases), (2) dans la conversion de la méthionine en cystéine et du tryptophane en niacine, (3) dans la néoglucogenèse comme coenzyme du glycogène phosphorylase ou encore (4) dans le métabolisme lipidique comme composante essentielle des enzymes facilitant la biosynthèse des sphingolipides.
Cette vitamine fut découverte dans les années 30, plus tardivement que la majorité des autres vitamines, sa carence ne causant pas de graves maladies bien que provoquant des
La pyridoxine est présente notamment dans la luzerne déshydratée, la levure de brasserie, le tourteau de soja et le son de riz (Sauvant et al., 2004).
2.1.5. Folates (vitamine B9)
Le nom « folates » est un terme générique employé pour désigner l’acide folique (ex. acide ptéroylmonoglutamique) et ses dérivées ayant la même activité biologique. L’acide folique est composé de trois éléments principaux : (1) un noyau ptéridine, (2) une molécule d’acide para-aminobenzoïque et (3) une molécule d’acide glutamique. L’ajout de plusieurs acides glutamiques permet de former des polyglutamates. Les dérivés réduits des polyglutamates (ex. dihydrofoliques ou tétrahydrofoliques) sont des accepteurs de groupements monocarbonés (Le Grusse et Watier, 1993; Figure 2.6).
La détermination de la structure de l’acide folique et sa synthèse furent réalisées en 1945. Durant la même année, on découvrit que cette vitamine permettait de soigner l’anémie mégaloblastique de la grossesse.
L’acide folique est essentiel à la synthèse des purines et pyrimidines pour la formation de l’ADN lors de la division cellulaire. Il joue un rôle clé dans la synthèse de novo de groupements méthyles (Girard et Matte, 2006). Ces groupements sont des constituants de la choline et des phospholipides nécessaires au maintien de l’intégrité des membranes cellulaires et à la formation des phospholipides sécrétés dans le lait chez la vache.
Les sources végétales d’acide folique pour les ruminants sont les drêches de brasserie, la farine de luzerne, les graines de soja et le tourteau d’arachide (Sauvant et al., 2004).
Acide folique
Acide dihydrofolique
Acide tétrahydrofolique
Polyglutamates
Figure 2.6 : Structure chimique de l’acide folique et de ses dérivés.
2.1.6. Vitamine B12
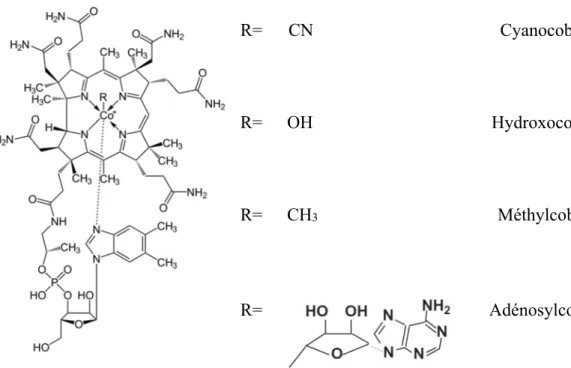
Le nom vitamine B12 est un terme générique regroupant la cyanocobalamine (-CN), l'hydroxocobalamine (OH-), la méthylcobalamine (-CH3), et l’adénosylcobalamine (5’-désoxyadénosine; Figure 2.7). Ces molécules ont la particularité d’être des corrinoïdes (c’est-à-dire composés d’un noyau corrine) possédant la même activité biologique (Combs Jr., 2012).
R= CN Cyanocobalamine
R= OH Hydroxocobalamine
R= CH3 Méthylcobalamine
R= Adénosylcobalamine
Figure 2.7 : Structure chimique de la cobalamine et de ses dérivés.
La vitamine B12 fut la dernière vitamine à être découverte en 1945. Cette vitamine possède un cobalt dans sa molécule complexe et est la plus lourde des vitamines. Deux enzymes sont vitamine B12-dépendantes. La première est la méthionine synthase, essentielle pour le transfert d'une unité de carbone à partir de la forme méthylée de l'acide folique à un acide aminé, l'homocystéine, le tout afin de former la méthionine. La seconde enzyme est la méthylmalonyl-CoA mutase qui transforme le méthylmalonyl-CoA en succinyl-CoA. Le méthylmalonyl-CoA provient de la dégradation des acides gras à nombre impair de carbones, dont le propionate et certains acides aminés (Girard et Matte, 2006).
La vitamine B12 est la seule vitamine à être synthétisée uniquement par les microorganismes, par conséquent, il est peu probable de trouver cette vitamine dans les aliments (McDowell, 2000). Cette vitamine est trouvée dans la levure de brasserie et dans les produits d’origine animale comme la farine de poisson et la poudre de lait entier (Sauvant et al., 2004).
Synthèse ruminale
La quantité de vitamines disponibles pour la vache laitière dépend de la consommation de vitamines, de la synthèse et de la dégradation de la vitamine dans le rumen et de sa biodisponibilité.
La synthèse apparente des vitamines B dans le rumen est le résultat de la différence entre la quantité ingérée (la teneur en vitamines de la ration multipliée par la consommation de matière sèche) et le flux total de vitamines mesuré au duodénum. L’expression «apparente » se rapporte à la quantité de vitamines mesurée sans prendre en compte les molécules éventuellement détruites dans le rumen, absorbées à travers les parois du rumen ou encore, dégradées par les microorganismes pour leur métabolisme.
Plusieurs facteurs peuvent influencer la production de vitamines B dans le rumen et donc, possiblement les quantités de vitamines disponibles pour l’animal. Les principaux facteurs sont décrits dans les sections ci-dessous.
2.2.1. Rapport fourrage:concentré
Le rapport fourrage:concentré est un des facteurs alimentaires les plus étudiés chez les ruminants. En effet, il est connu pour altérer de manière substantielle les populations et l’activité de la microflore du rumen. Par conséquent, il peut potentiellement modifier la production ou la dégradation de vitamines B.
L’étude de Hunt et al. (1941) a montré que la concentration de riboflavine présente dans le contenu du rumen de bouvillons était plus élevée lorsqu’ils étaient nourris avec une ration composée de foin de luzerne, de maïs et d’un supplément protéique, plutôt qu’avec une ration composée exclusivement de foin. Quelques années plus tard, Hayes et al. (1966) ont observé qu’une ration composée exclusivement de concentrés induisait des concentrations plus élevées de thiamine, de niacine, d’acide pantothénique, d’acide folique et de vitamine B12 dans la fraction liquide du contenu du rumen en comparaison avec une ration composée de maïs et de foin.
Des études plus récentes ont rapporté que l’augmentation de la proportion de fourrage de 35 à 60% entraînait une diminution de la synthèse apparente de vitamine B12 dans le rumen en fonction de la matière organique ingérée (Schwab et al., 2006).
2.2.2. Niveau d’ingestion
Au début des années 70, Sutton et Elliot (1972) ont démontré que la production de vitamine B12 biologiquement active augmentait avec le niveau d’ingestion de matière sèche digestible chez les moutons. Par contre, le niveau de vitamine B12 totale n’était pas influencé par le niveau d’ingestion, indiquant une réduction relative dans la synthèse des analogues de la vitamine B12, molécules semblables à la vitamine B12 mais dépourvues d’activité biologique chez l’animal.
Des effets similaires ont été trouvés par Zinn et al. (1987) chez les bouvillons. Ils ont observé une hausse de la synthèse apparente au rumen de thiamine, niacine, vitamine B6 et vitamine B12 lorsque l’ingestion de matière organique digestible augmentait.
2.2.3. Taille des particules
Les études des effets de la taille des particules sur la synthèse des vitamines B sont rares. Dans les années 40, Hunt et al. (1943) ont observé chez le bouvillon que le maïs moulu induisait une plus grande concentration de riboflavine dans le contenu du rumen que des grains de maïs entiers. Quelques années plus tard, dans une autre étude consacrée à ce sujet, Hayes et al. (1966) ont confirmé que les concentrations de thiamine, riboflavine, niacine et vitamine B6, étaient plus élevées dans la fraction liquide du contenu du rumen de bouvillons recevant du maïs moulu plutôt que floconné. Cette étude a aussi mis en évidence que la concentration de riboflavine était supérieure dans le contenu du rumen des animaux alimentés avec le foin moulu plutôt qu’avec le foin long.
2.2.4. Source de glucides et niveau d’azote
Afin de dégrader les glucides dans le rumen, la population microbienne procède à l’hydrolyse extracellulaire des polymères glucidiques puis à la fermentation intracellulaire des glucides simples. La dégradation de l’azote, quant à elle, liée à l’activité des protéases
microbiennes, dépend de la microflore du rumen et du type de protéines ingérées. Le synchronisme énergie-protéine est un aspect important afin de maximiser le métabolisme microbien (NRC, 2001). L’influence des apports en azote et en glucides sur la synthèse des vitamines B a été évaluée par Lardinois et al. (1944). Ces auteurs ont démontré que l’ajout d’urée augmentait la synthèse de riboflavine, d’acide nicotinique, de biotine et d’acide pantothénique, cet effet étant plus marqué lorsque le niveau de glucides facilement fermentescibles était plus élevé. Plus récemment, Beaudet et al. (2016) ont rapporté qu’une diminution de l’apport alimentaire d’azote réduit la synthèse apparente de vitamine B6 et de folates dans le rumen. Même si les diètes étaient isoénergétiques, remplacer l’amidon par des sources de fibres a diminué la synthèse apparente au rumen de riboflavine, de vitamine B6 et de folates mais a augmenté la synthèse de la vitamine B12.
2.2.5. Les additifs et le cobalt
L’utilisation d’additifs peut modifier les conditions du rumen (i.e.: pH, osmolarité) et, par conséquent, affecter la population microbienne en modifiant tant le nombre de bactéries que les proportions des différentes espèces ou leur activité. Celles-ci étant responsables de la synthèse et de la dégradation des vitamines B, l’état de la population microbienne influence directement la production de ces vitamines. Teeri et al. (1950) ont observé que des génisses nourries au foin supplémenté en sulfathalidine (qui cause une réduction de l’activité de synthèse de la flore intestinale) présentaient des concentrations urinaire et fécale de thiamine inférieures par rapport à des individus qui recevaient des diètes non supplémentées. Cependant, cet effet n’a pas été observé pour la niacine, la riboflavine et l’acide pantothénique.
Kon et Porter (1953) ont noté qu’une supplémentation journalière de 200 mg de pénicilline-procaïne à la ration de bouvillons n’avait aucun effet sur les concentrations de riboflavine, de pyridoxine, de biotine et de vitamine B12 dans le rumen. Quelques années plus tard, Hayes et al. (1966) ont constaté la même absence d’effet de l’incorporation d’un mélange d’antibiotiques sur les concentrations de thiamine, de riboflavine, de biotine, de niacine, d’acide folique, d’acide pantothénique et de vitamine B12 dans le rumen de
Des bouvillons recevant des rations supplémentées avec 0,5 g de chlorotétracycline pendant 15 jours ont présenté une réduction de la teneur en riboflavine et une augmentation des teneurs en niacine et en acide pantothénique dans le contenu du rumen (Chance et al., 1953). Cependant, Miller et al. (1986), bien qu’utilisant le même antibiotique à une dose de 7,8 mg/kg de ration ou 70 mg par animal, n’ont trouvé aucun effet sur le flux duodénal des vitamines du complexe B chez des bouvillons. Par contre, dans cette dernière étude, les auteurs ont constaté que l’ajout de monensin dans la ration diminuait le flux duodénal de niacine, augmentait le flux duodénal de riboflavine, mais n’avait aucun effet sur les flux duodénaux de thiamine et de biotine.
Le cobalt a fait l'objet de plusieurs études visant à augmenter la synthèse de la vitamine B12 dans le rumen. Kawashima et al. (1997) ont démontré que la production de vitamine B12 et de ses analogues était supérieure dans les cultures in vitro supplémentées avec différentes sources de cobalt. La supplémentation en cobalt chez le bovin augmentait aussi les concentrations de vitamine B12 dans le foie comparativement aux animaux non traités (Judson et al., 1997; Stangl et al., 2000). L’étude de Kincaid et Socha (2007) a démontré qu’une supplémentation en cobalt à la fin de la gestation et au début de la lactation pouvait augmenter la synthèse de la vitamine B12 dans le rumen, tel qu’indiqué par une tendance à l’augmentation des concentrations de vitamine B12 dans le colostrum et le lait chez les vaches recevant une ration supplémentée.
Supplémentation en vitamines B
Comme cela a déjà été mentionné, les études réalisées dans les années 50 rapportant l’absence de symptômes de carences vitaminiques chez les ruminants adultes ne permettent pas de conclure que ces animaux n’ont pas en certaines occasions besoin d’une supplémentation en vitamines B. En effet, l’efficacité métabolique peut grandement être affectée lorsque les apports en vitamines B sont insuffisants et la mise sur le marché de suppléments vitaminiques protégés de la dégradation dans le rumen semble pouvoir pallier à ces manques (Sacadura et al., 2008). Cependant, la réponse à ces suppléments, protégés ou non de la dégradation dans le rumen, est variable.
Cette variabilité est mise en évidence lorsqu’on compare les résultats des nombreuses études sur les effets de la supplémentation en niacine sur les performances de production des vaches laitières. Par exemple, plusieurs études rapportent qu’une supplémentation journalière de 6 mg pendant la période de transition et le début de la lactation augmente la production de lait (Kung et al., 1980; Riddel et al., 1981; Dufva et al., 1983; Muller et al., 1986; Jaster and Ward, 1990). En outre, cette plus grande production de lait lorsque les vaches reçoivent un supplément de niacine semble être due à l’augmentation de l’ingestion de matière sèche. Par ailleurs, Cervantes et al. (1996) ont rapporté une augmentation de la quantité totale de protéines produites quotidiennement dans le lait lorsque 12 g de nicotinamide ont été fournis aux vaches à environ 122 jours de lactation. Fronk et Schultz (1979) ont rapporté des effets positifs d’une supplémentation journalière de 12 g de niacine pendant 7 jours sur la santé des vaches souffrant de cétose clinique ou sous-clinique. De plus, cette supplémentation augmente la production de protéine microbienne et de propionate dans le rumen (Riddell et al., 1980; Horner et al., 1988). Cependant, d’autres études n’ont observé aucun effet de la supplémentation en niacine que ce soit sur les performances laitières (Driver et al., 1990; Minor et al., 1996), les concentrations plasmatiques de corps cétoniques ou d’acides gras non-estérifiés (Skaar et al., 1989; Bernard et al., 1995), ou encore, sur la fermentation microbienne dans le rumen (Kung et al., 1980; Campbell et al., 1994).
La variabilité des réponses suite à l’utilisation de suppléments de vitamines a aussi été rapportée avec d’autres vitamines (Girard et Matte, 1998; Shaver et Bal, 2000; Girard et al., 2005; Preynat et al., 2009; Evans et Mair, 2013 ). Ces divergences pourraient être liées au stade de lactation, au statut métabolique de la vache, aux caractéristiques de la diète, ou bien au métabolisme des microorganismes dans le rumen. Cependant, les connaissances des facteurs régulant ce dernier, particulièrement chez la vache laitière, sont presque inexistantes (Seck et al., 2012).
Ce projet devrait permettre d’identifier les facteurs critiques affectant la synthèse et la disponibilité des vitamines B pour la vache. La banque de données formées à partir de la composition chimique des aliments incluant leurs teneurs en vitamines B, de la synthèse
nutritionnelles permettra le développement d’un modèle de prédiction des quantités de vitamines B disponibles pour la vache en fonction de la régie d’alimentation. Ce modèle permettra aux producteurs d’ajuster les rations en fonction des besoins de la vache et, le cas échéant, d’évaluer la pertinence d’utiliser des suppléments de vitamines.
Analyse des vitamines B
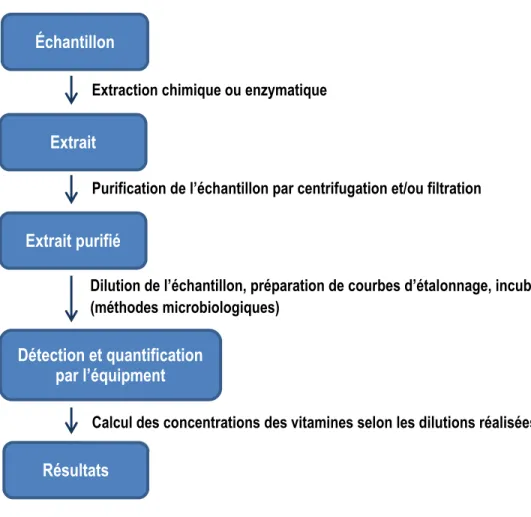
Les concentrations des vitamines dans les aliments et autres substrats peuvent être déterminées par des méthodes biologiques, microbiologiques et physicochimiques. Les méthodes biologiques permettent de donner des réponses qualitatives; elles consistent à tester si un aliment, lorsqu’offert aux animaux, permet de soigner des symptômes associés à une carence en une vitamine, suggérant la présence d’une quantité significative de celle-ci dans l’aliment. Les méthodes microbiologiques et physicochimiques, réalisées en plusieurs étapes permettent d’obtenir des réponses quantitatives (Figure 2.8).
Figure 2.8. Représentation schématique de l’analyse des vitamines. Extraction chimique ou enzymatique
Purification de l’échantillon par centrifugation et/ou filtration
Dilution de l’échantillon, préparation de courbes d’étalonnage, incubation (méthodes microbiologiques) Échantillon Extrait Extrait purifié Détection et quantification par l’équipment Résultats
La détermination des concentrations en vitamines des échantillons débute par l’extraction de ces molécules de la matrice. La méthode d’extraction varie selon la vitamine analysée, la nature de la matrice et la résistance de la vitamine à l’égard de la chaleur, de la lumière et des valeurs extrêmes de pH. Il est essentiel que les vitamines soient extraites de la matrice sous une forme permettant une détermination précise au moyen de la technique d’analyse utilisée. Une procédure d’extraction efficace homogénéise et concentre l’échantillon, isole le composé de sa liaison avec la protéine et élimine autant que possible les substances interférentes connues (Ball, 2006). Les méthodes d’analyse de vitamines utilisées dans cette thèse sont (1) la chromatographie en phase liquide à haute performance (CLHP), (2) la méthode microbiologique et (3) la méthode radio-isotopique. Celles-ci sont décrites en détails au Chapitre 3 de ce manuscrit. Des informations complémentaires à ces méthodes suivent.
2.4.1. Chromatographie en phase liquide à haute performance (CLHP)
Les concentrations de thiamine, riboflavine, niacine et vitamine B6 ont été déterminées grâce à la CLHP qui permet une détection très précise, rapide et complète. Cette méthode consiste à appliquer un petit volume d’extrait stable d’échantillon (10 [thiamine] – 20 [riboflavine, niacine et vitamine B6] µL) dans une colonne remplie d’un matériel microparticulé (5µm - phase stationnaire) dont les composés sont élués sous haute pression avec un liquide (phase mobile). La détection de ces composés se fait à partir de leur séparation et d’une surveillance continue de la fluorescence de l’effluent de la colonne par un détecteur fluorimétrique. L’objectif de cette séparation est d’obtenir des résolutions de pics de concentrations du composé le plus rapidement possible (Figure 2.9), celles-ci variant selon le composé, la pression et la composition de la phase mobile ainsi que la phase stationnaire et la composition interne de la colonne.
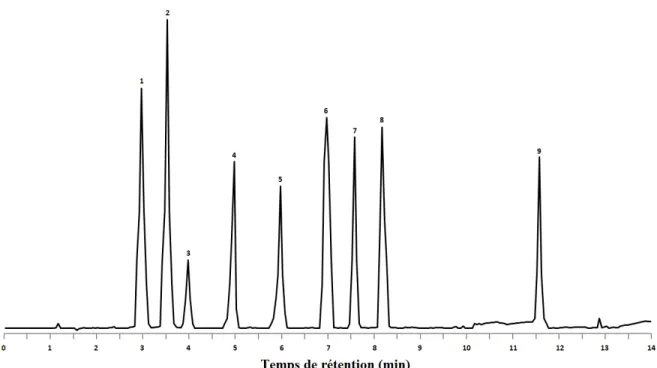
Figure 2.9. Exemple théorique d’un chromatogramme produit par une méthode d’analyse de vitamines par CLHP en phase inverse.
Piques : (1) acide nicotinique; (2) flavine adénine dinucléotide; (3) thiamine; (4) flavine mononucléotide; (5) pyridoxamine; (6) pyridoxal; (7) riboflavine; (8) pyridoxine; (9) nicotinamide.
Les analyses des vitamines par CLHP ont été réalisées en duplicata et les mesures de variations intra- et inter-essai ainsi que la mesure de leur spécificité (e.g. : pourcentage de recouvrement) sont présentées dans le Tableau 2.1. Les contrôles utilisés pour calculer ces variations ont été des échantillons de digesta duodénal et de ration totale mélangée (RTM).
Tableau 2.1. Reproductibilité inter- et intra-essai et pourcentage de recouvrement
Inter-essai Intra-essai Recouvrement (%)
Duodénal RTM Duodénal RTM Duodénal RTM
Thiamine 11,0 8,0 5,8 1,1 99,2 91,3
Riboflavine 15,4 14,3 3,5 3,8 101,0 94,3
Niacine 12,1 9,5 1,8 5,5 89,1 88,0
2.4.2. Méthode microbiologique pour la détermination des folates
La méthode microbiologique utilisée a été basée sur la croissance du Lactobacillus
rhamnosus, une bactérie dépendante de l’apport de folates parmi ses plusieurs formes
(acide folique, tétrahydrofolate, méthyltetrahydrofolate, etc.), ce qui justifie l’utilisation de la conjugase lors de l’extraction). L’acide folique a été extrait d’un échantillon et déterminé par un test commercial utilisant une plaque de microtitration (VitaFast, Folic Acid, R-Biopharm Inc.). La croissance de ce microorganisme a été mesurée par la turbidité en suspension et comparée à la courbe d'étalonnage (Figure 2.10) par un spectrophotomètre. Les coefficients de variations intra- et inter-essai ont été respectivement de 3,55 et 13,78 % pour les échantillons de RTM.
2.4.3. Méthode de radio-dosage pour la détermination de vitamine B12
La méthode de dosage radio-isotopique est basée sur le principe de la compétition entre la vitamine B12 non marquée et ses homologues marqués sur un nombre restreint de sites de fixation. Par conséquent, la plus grande présence du composé non marqué se traduit par un taux plus faible de radioactivité sur le facteur intrinsèque purifié (isolé de l’estomac du porc et ayant une forte affinité pour la vitamine). Les coefficients de variations intra- et inter-essai ont été respectivement de 3,16 et 4,46% pour le contrôle utilisant le digesta duodénal. Les coefficients de variations des échantillons de RTM n’ont pas été calculés du au fait que les concentrations de vitamines B12 sont très souvent sous le seuil de détection de la méthode pour ce type de matériel.
3. ARTICLE 1 :
ESPÈCE
FOURRAGÈRE /
FORAGE FAMILY
Effects of forage family on apparent ruminal synthesis of B vitamins
in lactating dairy cows
D. S. Castagnino,*†1 M. Seck,*†1 V. Beaudet,*† K. L. Kammes,‡ J. A. Voelker Linton,‡ M. S. Allen,‡ R. Gervais,† P. Y. Chouinard† and C. L. Girard*
*Sherbrooke Research and Development Centre, 2000 College, Sherbrooke, Québec, Canada, J1M 0C8
†Département des sciences animales, Université Laval, 2425 rue de l’Agriculture, Québec, Québec, Canada, G1V 0A6
‡Department of Animal Science, Michigan State University, East Lansing 48824-1225 1Both authors contributed equally to these studies
Cet article est publié dans la revue Journal of Dairy Science.
Référence : Castagnino D. S., M. Seck, V. Beaudet, K. L. Kammes, J. A. Voelker Linton, M. S. Allen, R. Gervais, P. Y. Chouinard and C. L. Girard. 2016. Effects of forage family on apparent ruminal synthesis of B vitamins in lactating dairy cows. J. Dairy Sci. 99, 1884-1894.
Résumé
Les effets de l’espèce fourragère (légumineuse vs graminée) sur la synthèse apparente dans le rumen (SAR) et les apports post-ruminaux de vitamines B ont été évalués au cours de deux expériences. Des rations contenant comme seul fourrage soit l’ensilage de luzerne (LU), soit l’ensilage de dactyle (DA) ont été offertes à des vaches Holstein en lactation munies de canules au rumen et au duodénum selon un plan d’expérience en chassé-croisé. Au cours de l’expérience 1 (E1) des rations à base de LU ou de DA (~23% de fibres insolubles dans le détergent neutre (NDF) provenant du fourrage et ~27% de NDF total dans la ration) ont été offertes à 8 vaches pendant deux périodes de 15 jours. Au cours de l’expérience 2 (E2) des rations à base de LU et de DA (~25% NDF provenant du fourrage et ~30% de NDF total dans la ration), ont été offertes à 13 vaches pendant deux périodes de 18 jours. Les concentrations de thiamine, riboflavine, niacine, vitamine B6, folates et vitamine B12 ont été déterminées dans des échantillons d’aliments et de digesta duodénaux. La SAR a été calculée sur une base journalière comme étant le flux duodénal (FD) mesuré pour chaque vitamine moins la quantité respective ingérée. L’espèce fourragère a affecté l’ingestion, la SAR et le FD des vitamines B. L’utilisation de luzerne dans les rations a entraîné une ingestion supérieure de vitamine B6 et inférieure de folates par rapport au dactyle. Dans E1, les ingestions de riboflavine et niacine ont été supérieures pour DA tandis que dans E2, l’ingestion de thiamine a été supérieure et celle de riboflavine a été inférieure pour DA comparativement à LU. Malgré la faible concentration de folates dans les deux ensilages, l’ingestion de folates a été supérieure pour DA, en raison des concentrations de tourteau de soja différentes entre les diètes. L’espèce fourragère n’a pas affecté la SAR de niacine et de folates. Le flux duodénal d’azote microbien était corrélé positivement avec la SAR de riboflavine, niacine, folates et vitamines B6 et B12 mais négativement avec la SAR de thiamine. Les SAR de folates et de vitamine B12 semblent être liées à l'activité de la biomasse microbienne. Les changements de la composition nutritionnelle des rations pourraient affecter les populations microbiennes dans le rumen et leur métabolisme des vitamines B.
Abstract
Effects of forage family (legume vs. grass) on apparent ruminal synthesis (ARS) and post-ruminal supply of B vitamins were evaluated in two experiments. Diets containing either alfalfa (AL) or orchardgrass (OG) silages as the sole forage were offered to ruminally and duodenally cannulated lactating Holstein cows in crossover design experiments. Experiment 1 compared diets containing AL and OG (~23% forage NDF and ~27% total NDF) offered to 8 cows in two 15-d treatment periods. Experiment 2 compared diets containing AL and OG (~25% forage NDF and ~30% total NDF) offered to 13 cows in two 18-d treatment periods. Thiamin, riboflavin, niacin, vitamin B6, folates and vitamin B12 were analyzed in feeds and duodenal digesta. Apparent ruminal synthesis was calculated as the duodenal flow of each vitamin minus its intake. Forage family affected B-vitamin intakes, duodenal flow and ARS. In both experiments, AL diets increased vitamin B6 and decreased folate intakes. In Experiment 1, riboflavin and niacin intakes were greater with the OG diet whereas in Experiment 2, thiamin intake was greater but riboflavin intake was smaller with the OG diet. In spite of the low contribution of either silage to the dietary folate content, folate intake was greater with OG diets than AL, due to the difference in soybean meal contribution between diets. Niacin and folate ARS were not affected by the forage family. Duodenal microbial nitrogen flow was positively correlated with ARS of riboflavin, niacin, vitamin B6, folates and vitamin B12 but tended to be negatively correlated with thiamin ARS. Apparent ruminal synthesis of folates and vitamin B12 appear to be related with microbial biomass activity. Changes in nutrient composition of the diets likely affected the microbial population in the rumen and their B-vitamin metabolism.
Introduction
Studies on B-vitamin requirements of cattle, conducted more than 60 years ago, concluded that mature ruminants with a functional rumen did not require B-vitamin supplements because the amounts of vitamins provided by the diet or synthesized by rumen microorganisms were sufficient to avoid apparition of deficiency symptoms (Bechdel et al., 1926; Kon and Porter, 1954). However, deficiency symptoms are the last stage of the deficiency; deficiency appears as soon as the supply is inferior to the needs leading to biochemical changes (Combs Jr., 2012). Reports of positive production responses to B-vitamin supplementation in high-producing dairy cows (Girard and Matte 1998; 2005; Schwab et al., 2005; Chen et al., 2011) suggest that, under some conditions, the need for B vitamins frequently exceeds the supply from the diet and the synthesis by rumen microbes, leading to sub-optimal milk production and metabolic efficiency. Shaver and Bal (2000) reported that supplementing dietary thiamin increased milk and component yields.
A meta-analysis showed that a niacin supplement increased milk fat and protein yields and tended to increase feed efficiency (Schwab et al., 2005). Supplementary folic acid increased milk production of cows in the second lactation or greater (Girard and Matte, 1998). Girard and Matte (2005) observed that, when compared to a supplement of folic acid alone, a combined supplement of folic acid and vitamin B12 increased milk and milk component yields in primiparous cows during early lactation. However, some studies observed no effect of vitamin supplements (Girard et al., 2005; Rosendo et al., 2004). A better understanding of dietary factors driving the fate of B vitamins in rumen of dairy cows is essential to define their B-vitamin requirements. There is only a limited number of studies looking at the effects of dietary factors such as forage-to-concentrate ratio or grain processing on production of B vitamins by the ruminal microflora (Conrad and Hibbs, 1954; Hayes et al., 1966; Breves et al., 1981; Abel et al., 2001), but to our knowledge no data are available on the effects of forage family. The objective of the present study was to describe the effect of forage family (legume or grass) on B-vitamin supply and apparent ruminal synthesis (ARS) in lactating dairy cows.
Materials and methods
Experimental procedures of the 2 studies were conducted on cows from the Michigan State University Dairy Cattle Teaching and Research Center, and approved by the Institutional Animal Care and Use Committee at Michigan State University (East Lansing, MI). Prior to calving, cows were cannulated ruminally (Bar Diamond Inc., Parma, ID) and duodenally with a gutter-type T cannula placed approximately 10 cm distal to the pylorus (Joy et al., 1997). Surgery was performed at the Department of Large Animal Clinical Science, College of Veterinary Medicine, Michigan State University. Experimental procedures, measurements and analyses were described in detail by Voelker Linton and Allen (2008, Experiment 1), and Kammes and Allen (2012, Experiment 2).
3.4.1. Experiment 1
3.4.1.1. Cows and treatments
Eight multiparous cows were assigned randomly to treatment sequence in a crossover design with a 14-d preliminary period followed by two 15-d treatment periods. The two treatments were diets containing either alfalfa (AL; Medicago sativa L.) silage harvested at early bud stage or orchardgrass (OG; Dactylis glomerata L.) silage harvested at early boot stage as sole forage (Table 3.1). As both diets were formulated to contain 27% NDF and 23% forage NDF (Voelker Linton and Allen, 2008), the forage-to-concentrate ratios were 53:47 and 48:52 for AL and OG, respectively. Cows were fed ad libitum a TMR served once daily (1100 h) at 110% of expected intake.
Table 3.1. Ingredients, nutrient composition and B-vitamin concentrations of the two treatment diets containing alfalfa silage (AL) or orchardgrass silage (OG) in experiments 1 and 21 Experiment 1 Experiment 2 Item AL OG AL OG Ingredient, % DM Alfalfa silage 53.02 - 59.93 - Orchardgrass silage - 47.94 - 42.75
Dry ground corn 36.3 40.3 33.6 36.6
Soybean meal (48% CP) 6.50 7.00 - 11.8
Vitamin & mineral mix6,7 4.20 4.20 2.50 3.39
SoyPlus®8 1.30 1.30 3.99 3.99 Blood meal 0.30 0.90 - - Limestone - 0.40 - 0.30 Urea - 0.20 - 1.20 Nutrient DM, % as fed 43.6 50.6 54.5 52.3 OM, % of DM 91.5 91.5 92.7 91.1 NDF, % of DM 26.7 27.5 29.2 30.2 Forage NDF, % of DM 22.5 23.0 25.3 24.9 Indigestible NDF, % of DM 14.8 7.90 14.8 8.24 Starch, % of DM 30.2 32.1 27.3 29.6 CP, % of DM 18.3 18.8 18.4 17.0 B-vitamin concentrations, mg/kg of DM Thiamin 2.38 2.31 2.89 2.20 Riboflavin 36.93 46.07 66.95 20.50 Niacin9 22.32 56.44 46.70 50.60 Vitamin B610 17.63 11.84 7.38 3.36 Folates 0.36 0.46 0.36 0.44 Vitamin B12 ND11 ND 0.001 0.002
1Adapted from Voelker Linton and Allen (2008) and Kammes and Allen (2012).
2DM, 30.6% (as fed); and OM 88.7%; NDF, 42.6%; indigestible NDF, 25.2%; potentially digestible NDF,
17.4%; starch, 4.0%, CP, 20.5% (DM. basis).
3DM, 43.5% (as fed); and OM 91.9%; NDF, 42.3%; indigestible NDF, 23.0%; potentially digestible NDF,
19.3%; starch, 1.87%, CP, 22.5% (DM. basis).
4DM, 35.3% (as fed); and OM 89.2%; NDF, 48.0%; indigestible NDF, 13.1%; potentially digestible NDF,
34.9%; starch, 2.3%, CP, 20.4% (DM. basis).
5DM, 33.7% (as fed); and OM 90.3%; NDF, 58.2%; indigestible NDF, 16.1%; potentially digestible NDF,
42.1%; starch, 1.37%, CP, 11.4% (DM. basis).
6Experiment 1 = Vitamin & mineral mix contained, on a DM basis, 11.7% dicalcium phosphate, 11.1%
trace-mineral premix, 8.8% sodium bicarbonate, 2.3% magnesium oxide, 134.3 kIU/kg of vitamin A, 35.53 kIU/kg of vitamin D, 895.5 kIU/kg of vitamin E, and 65.2% ground corn grain as a carrier.
7Experiment 2 = Vitamin & mineral mix contained, on a DM basis, 16.5% sodium bicarbonate, 14.2%
magnesium sulfate, 7.1% salt, 5.8% dicalcium phosphate, 2.4% trace-mineral premix, 0.4% vitamin A, 0.4% vitamin D, 0.2% vitamin E, and 53.1% dry ground corn as a carrier.
8West Central Soy, Ralston, IA.
9Sum of the molar concentrations of niacin and nicotinamide; expressed as niacin-equivalents.
10Sum of the molar concentrations of pyridoxamine, pyridoxal, and pyridoxine; expressed as
pyridoxine-equivalents.