Potable water production by membrane processes: membrane characterization using a series of bacterial strains
9
0
0
Texte intégral
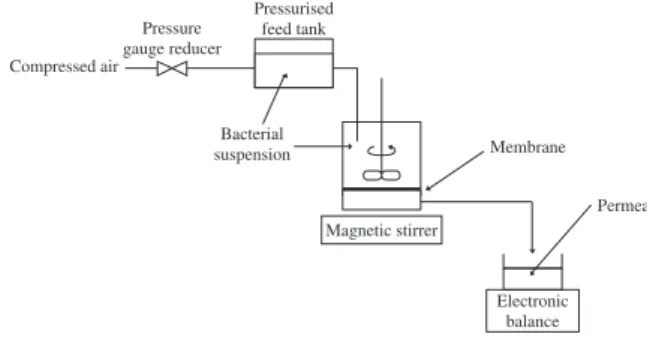
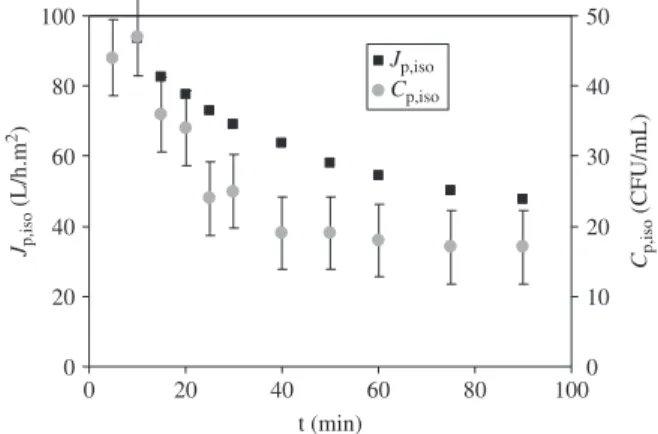
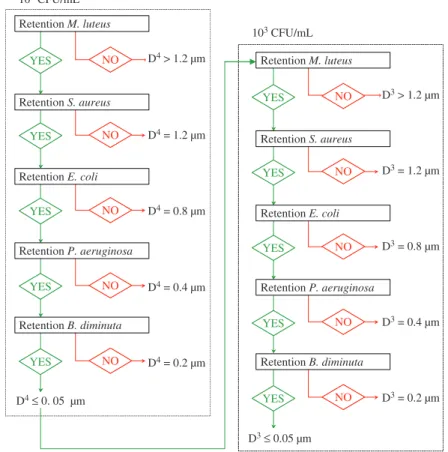
Figure
Documents relatifs
