SensoProTrainer®, the new fitness device : effects of a single bout of exercise and a four week training programme on motor memory consolidation
Texte intégral
Figure


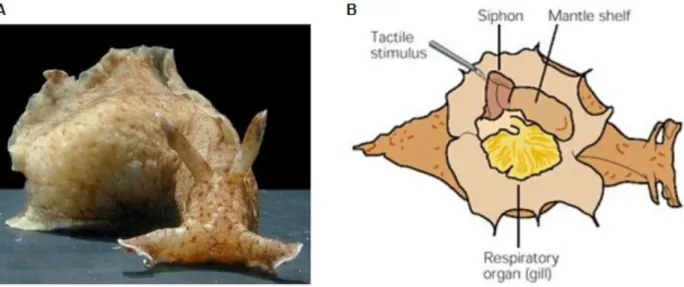
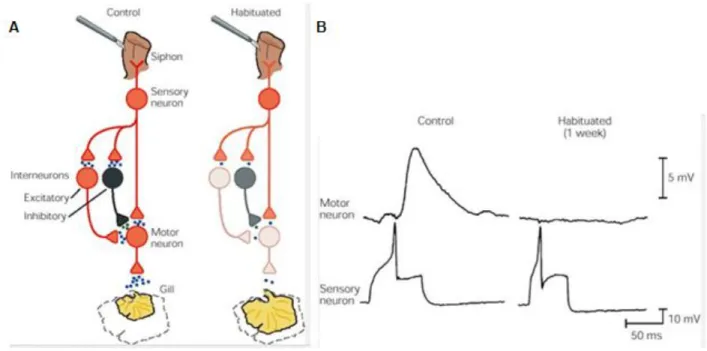
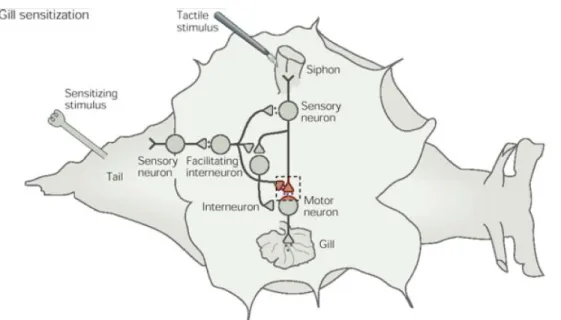
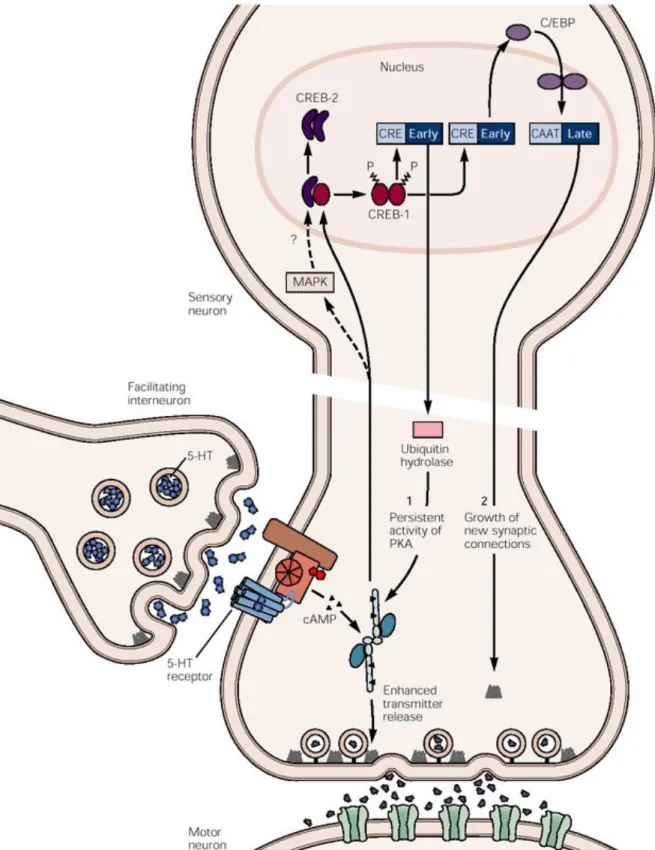
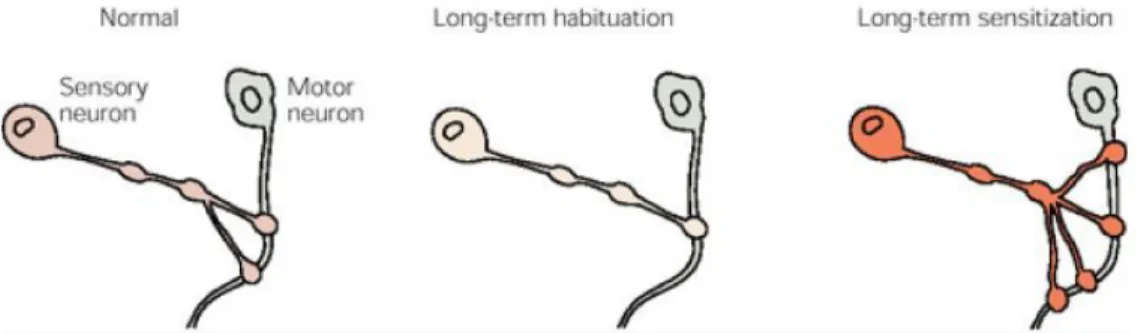
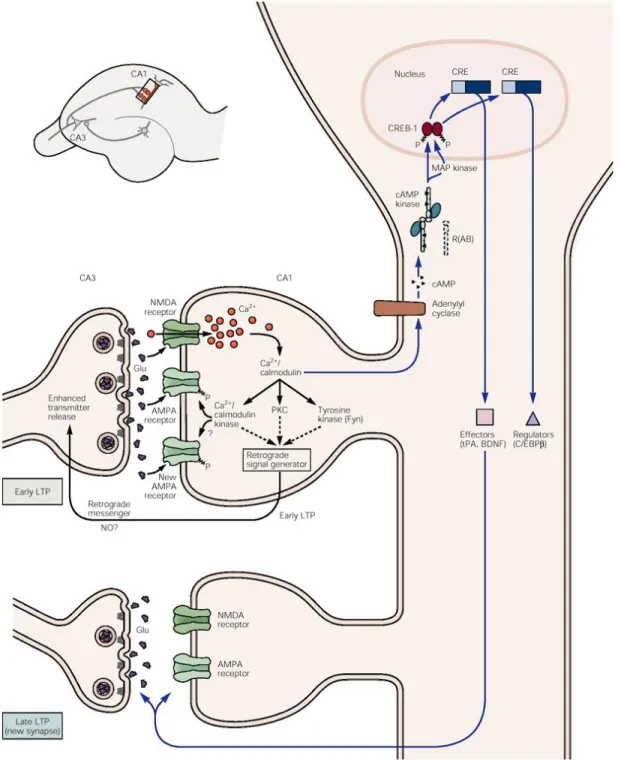
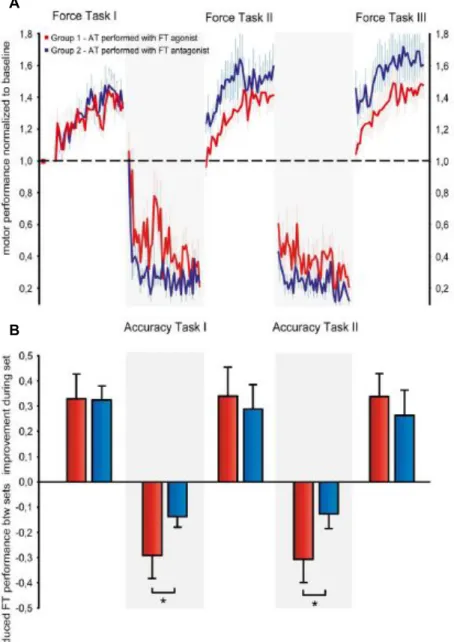
Documents relatifs
It shows that PhD students contribute to about a third of the publication output of the province, with doctoral students in the natural and medical sciences being present in a
Library science is no longer the major research focus in this field: the usage of the term “library science” and library-related words such as cataloging, reference, and
The High Court of Justice completely disregarded whether the construction of exclusive Israeli civil communities on an occupied land, over which the Israeli legal regime is
The second part is a meta-analysis of the effectiveness of gamification (assessed through RCT studies) applied to cognitive training with the following objectives: (1) assess
Quanti fication of epidermal DR5:GFP reflux signals (Figure 2F; see Supplemental Figure 1 online) con firmed slightly reduced basipetal re flux for twd1 (Bailly et al., 2008)
In the Falck case, the far-sighted family champion of change Alberto Falck—with crucial support of the external CEO Achille Colombo—was able to de-escalate the family business
[r]
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
