Cognitive and connectome properties detectable through individual differences in graphomotor organization
Texte intégral
Figure
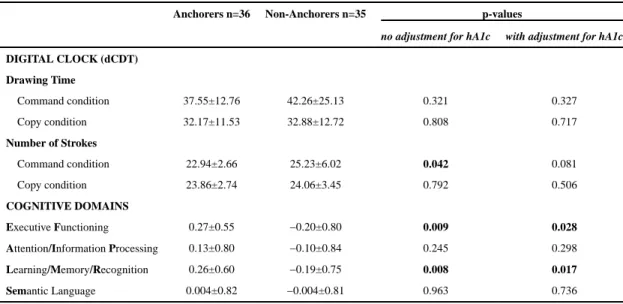
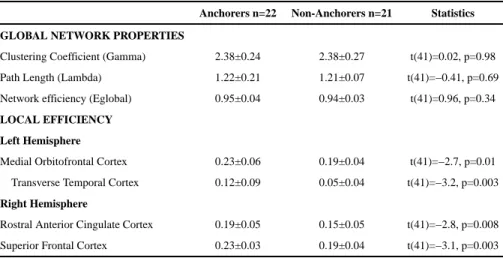
Documents relatifs
Thus, the AX-CPT is an interesting candidate task for a case study of the importance of considering psychometric properties in cognitive research related to individual differences..
Thus, individuals may endorse extreme pro-group actions to reaf fi rm their heritage cultural identity, and in doing so, realign balance in the friend- ship after perceiving
Post hoc analysis showed that across SL cue conditions, compared to control group the percentage change in SL variability was signi fi cantly reduced in PD patients in all
The associations of the Big Five traits, narcissism, and the other status update topics with the number of likes and comments received were examined on an exploratory basis to
As cognitive phenomena cannot be reduced to intra-individual processes (Forgas, 1981 a; b), and inversely, interactions are influenced by the representations held by group
The questionnaire included: socioeconomic characteristics (gender, birth date, father’s occupation, parents’ education, nationality, family structure, and family
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
To answer the research question, we followed a two-step between- subject experimental approach. In the first step, we performed an exploratory study, investigating whether and how
