Voltage- and cold-dependent gating of single TRPM8 ion channels
Texte intégral
Figure
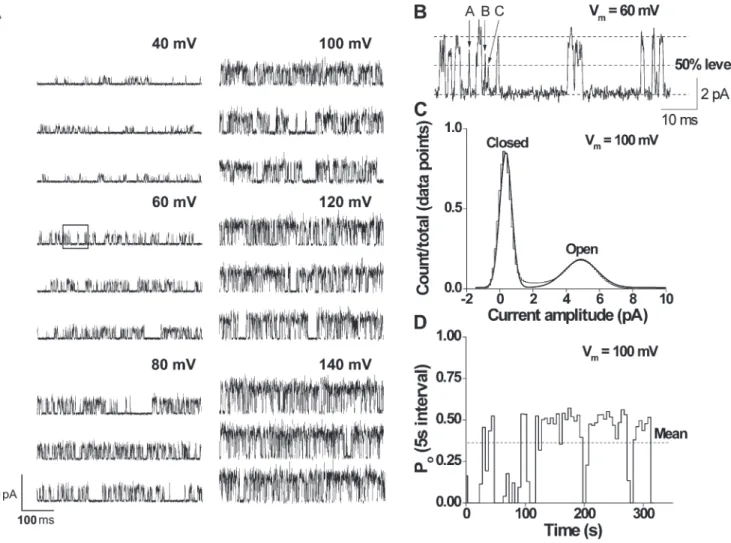
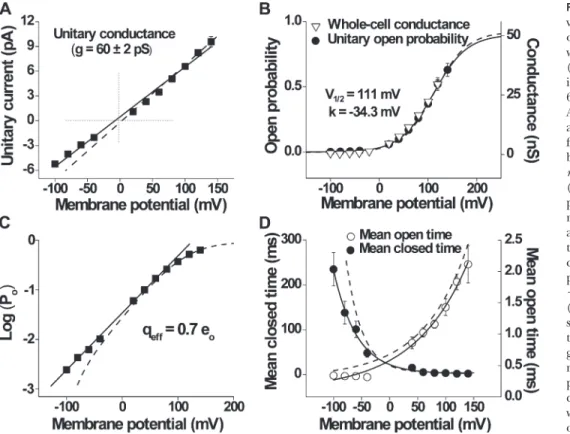
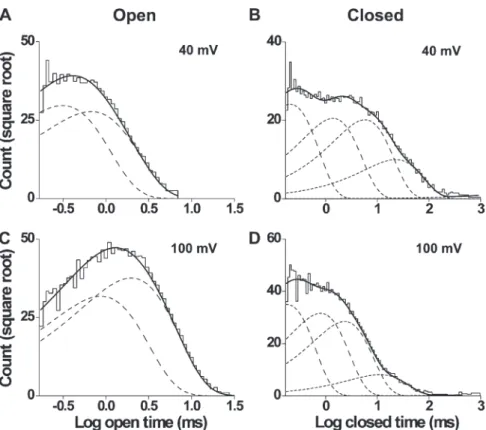

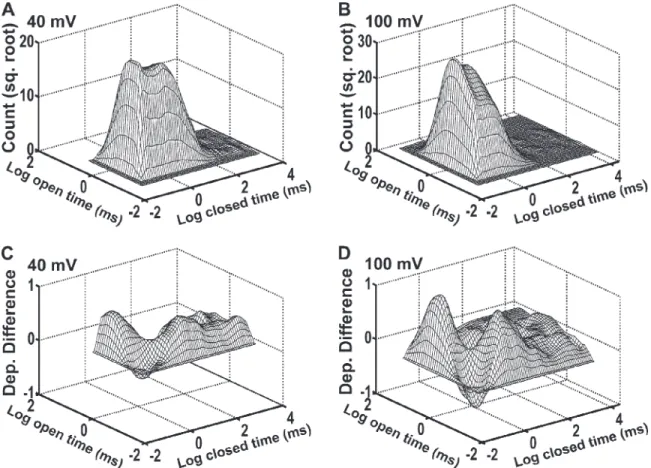
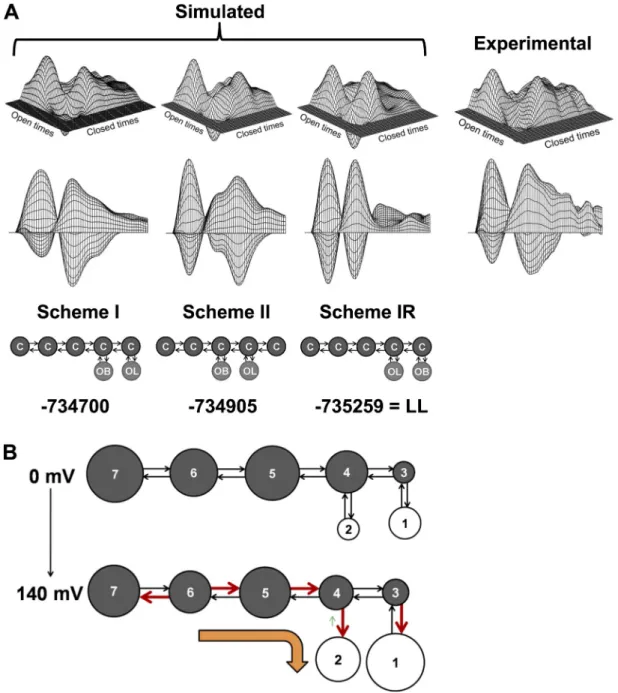
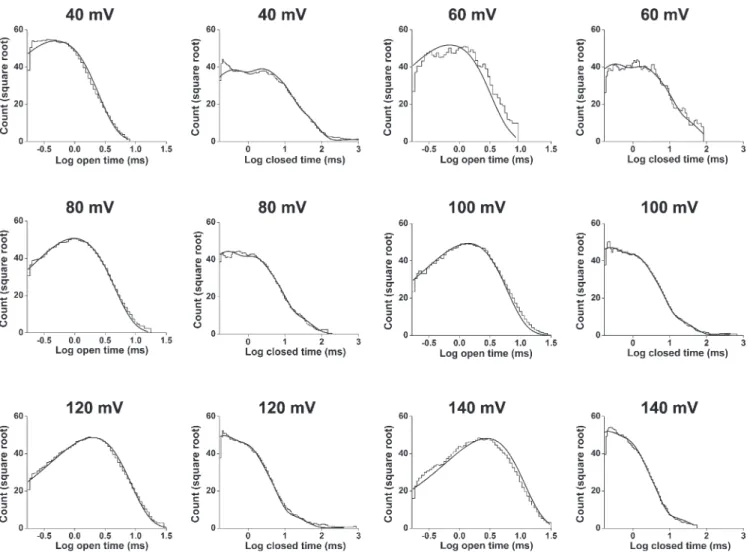
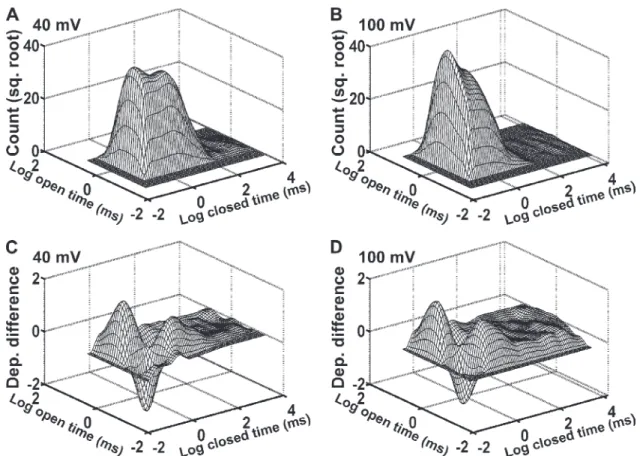
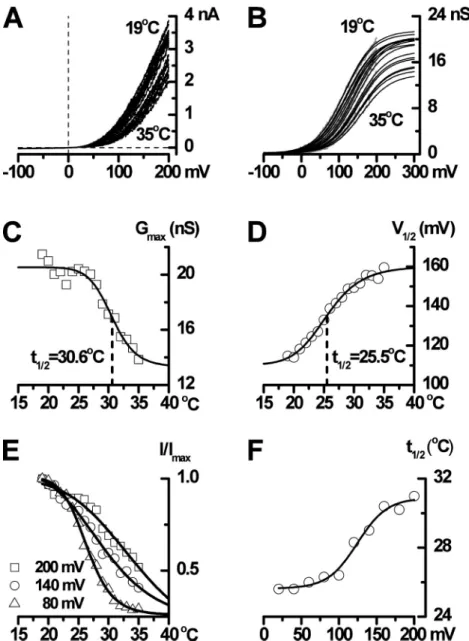
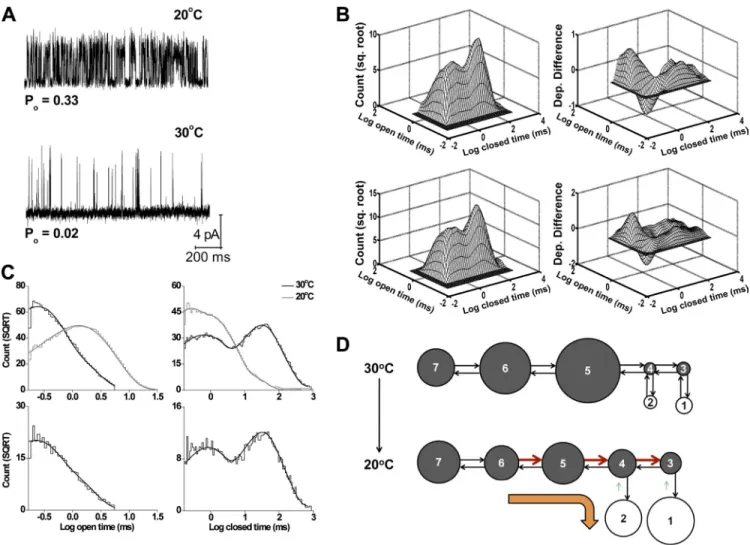
Documents relatifs
Effects of skin cooling on slow adapting type 1 (SA1) discharge and SA1 nerve fiber conduction: (a) Comparison of the SA1 discharge at 22 " C between wild- type (WT) and
To take into account this fact in our model, we adjusted the parameters to fit the quantity 0.2 × ( I + R ) from the model, with the total number of infected people (data in Table
[157] Vesikari, T., et al., Immunogenicity and safety of an investigational multicomponent, recombinant, meningococcal serogroup B vaccine (4CMenB) administered
The UBAIM input is taken from the neutral atmosphere models SOCRATES [Chabrillat, 2001] and MSISE [Hedin, 1991], the solar flux model SOLAR2000 [Tobiska et al., 2000], and from
L’objectif de ce travail consiste à évaluer les concentrations en plomb d’origine routière et d’établir une cartographie de la pollution à l’aide d’une espèce de
D’un autre côte, deux techniques récentes ont été utilisé pour confirmer l’existence du chaos dans le système étudié : le fer à cheval topologique, une méthode
Next, we used the conventional approach proposed by Pauly and Christensen (1995a) to estimate mean SPPRs of these species but using the mean transfer efficiency of 11.9%
Cells with higher acoustic contrast factor reach the nodal plane faster and can be collected in the central outlet while cells with a lower acoustic contrast factor stay on
