Tripartite siRNA Micelles as Controlled Delivery Systems for Primary Dendritic Cells
Texte intégral
Figure

Documents relatifs
Such diffusion processes capture both geometric and photometric information and give rise to local and global dif- fusion geometry (heat kernels and diffusion distances), which can
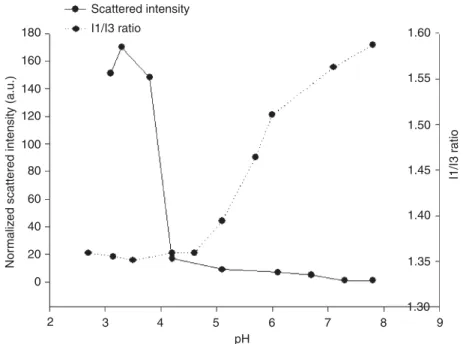
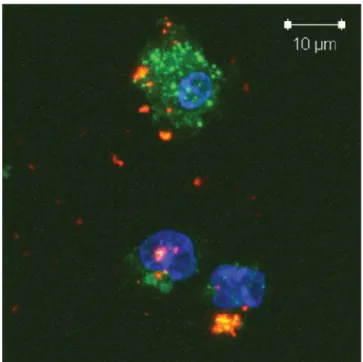
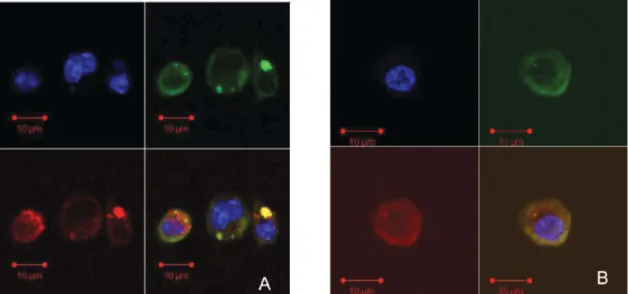
The micelles formed by PEG-PEI-PE possess several key features for siRNA and drug delivery, including (i) stability in physiological fluids thanks to the outer
F.P. designed and carried out the experiments. supervised the project. analysed the data and wrote the manuscript. gave technical support and conceptual advice. performed the ex
The effects of these changes on the physical properties (water absorption and linear shrinkage) and mechanical properties (flexural and compressive strengths) of
We have asked our subjects first to check certain traits that characterize a person presented in a sketch, and then to indicate the basis for doing so--what in the
Such fractional equations of order 2α exhibit new phenomena that do not have any counterpart in the theory of second order elliptic equations in divergence form: In [ 14 ], building
From this investigation, the AC of type 1 appears as a prevalent and conserved structural element exclusively associated to β-helical AT passenger and should promote further
Based on these observations we concluded that the presented reduction methods constitute e ffective, efficient, and scalable pre-processing steps toward the analysis of
