biodivMapR: An r package for α‐ and β‐diversity mapping using remotely sensed images
Texte intégral
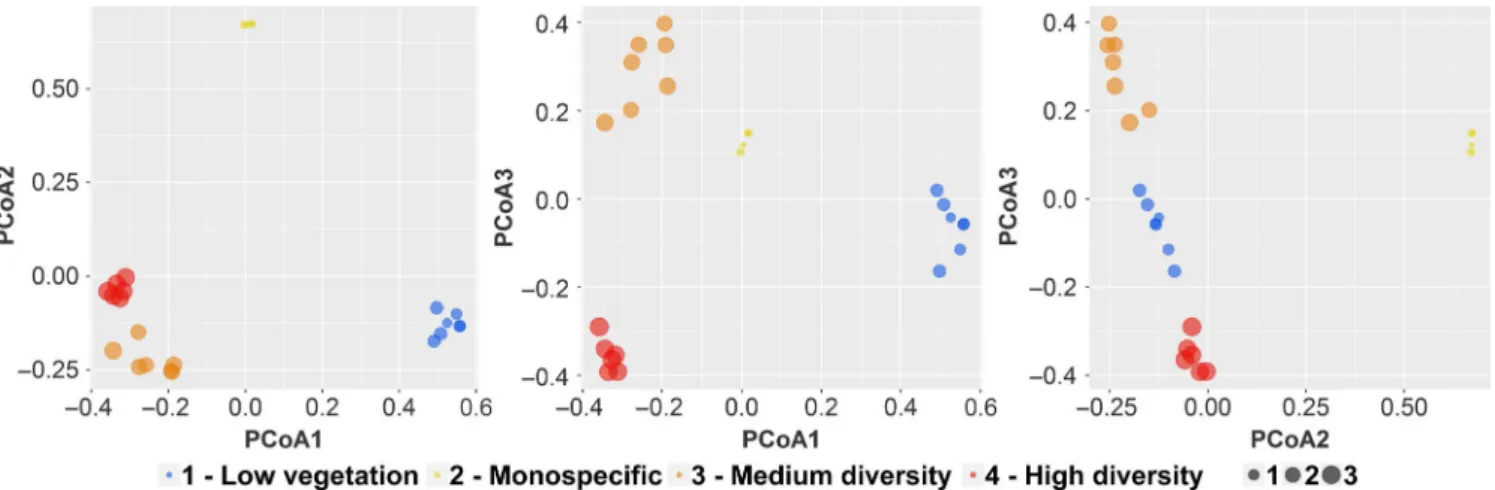
Figure
Documents relatifs
The same conclusions than in Section 4.2 can be driven: some metabolites were extracted by both analyses (creatinine, betaine, hippuric acid, guanidinoacetic acid, alanine,
The ZIP employ two different process : a binary distribution that generate structural
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
The problems as well as the objective of this realized work are divided in three parts: Extractions of the parameters of wavelets from the ultrasonic echo of the detected defect -
The problems as well as the objective of this realized work are divided in three parts: Extractions of the parameters of wavelets from the ultrasonic echo of the detected defect -
Figure 6 shows that the packages Re- moteDirectory and TelNetWordNet are referencing together the same set of packages within Network: both refer to classes into Kernel, Protocols
The ARSIS concept (from its French name Amélioration de la Résolution Spatiale par Injection de Structures) was designed to improve the spatial resolution of images in a set
In practice, RPCA via L+S decomposition is suitable for video processing tasks in which (1) the observed video can be viewed as the sum of a low-rank clean video without
