Towards reconciling structure and function in the nuclear pore complex
12
0
0
Texte intégral
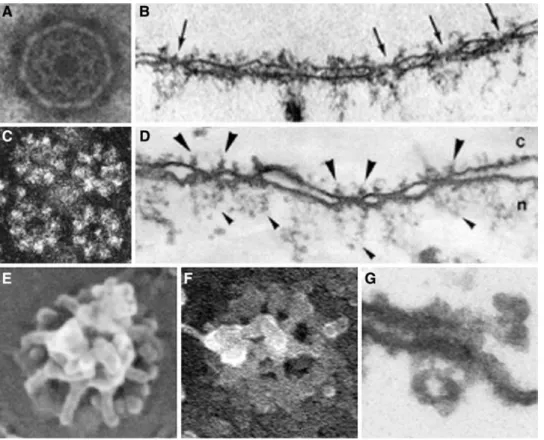
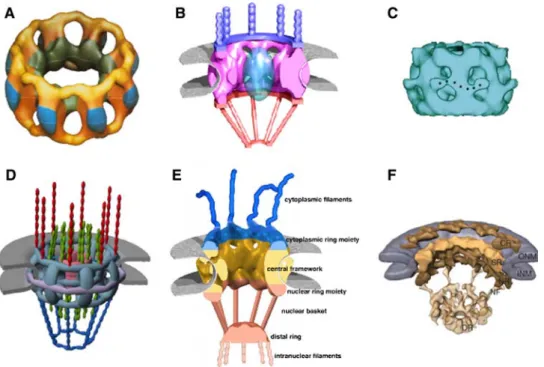
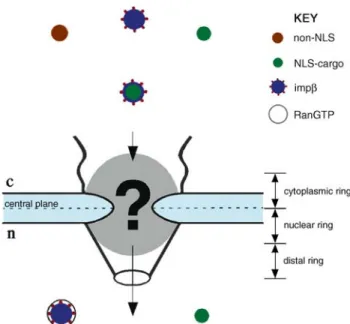
Figure

Documents relatifs
