THE B O T A N I C A L
R E V I E W
VOL. 66
JANUARY-MARCH
2000
NO. 1
A Revised Classification of the Apocynaceae s.l.
M A R Y E . E N D R E S SInstitute of Systematic Botany University of Zurich 8008 Zurich, Switzerland
A N D
P E T E R V . B R U Y N S
Bolus Herbarium University of Cape Town Rondebosch 7700, South Africa
I. AbstractYZusammen fassung . . . 2
II. Introduction . . . 2
III. D i s c u s s i o n . . . 3
A. Infrafamilial Classification o f the A p o c y n a c e a e s.str . . . 3
B. R e c o g n i t i o n o f the F a m i l y Periplocaceae . . . 8
C. Infrafamilial Classification o f the A s c l e p i a d a c e a e s.str . . . 15
1. R e c o g n i t i o n o f the S e c a m o n o i d e a e . . . 15
2. Relationships within the A s c l e p i a d o i d e a e . . . 17
D. Coronas within the A p o c y n a c e a e s.l.: H o m o l o g i e s and Interpretations . . . 22
IV. Conclusion: The A p o c y n a c e a e s.1 . . . 27
V. T a x o n o m i c T r e a t m e n t . . . 31
A. Key to the S u b f a m i l i e s o f the A p o c y n a c e a e s.1 . . . 31
1. R a u v o l f i o i d e a e Kostel . . . 32 a. A l s t o n i e a e G. Don . . . 33 b. V i n c e a e D u b y . . . 34 c. W i l l u g h b e e a e A. DC . . . 34 d. T a b e r n a e m o n t a n e a e G. Don . . . 35 e. M e l o d i n e a e G. Don . . . 36 f. Hunterieae Miers . . . 36 g. P l u m e r i e a e E. M e y . . . 36 h. Carisseae D u m o r t . . . 37 i. A l y x i e a e G. Don . . . 37 C o p i e s o f t h i s i s s u e [66(1)] m a y b e p u r c h a s e d f r o m the N Y B G Press, The N e w Y o r k B o t a n i c a l G a r d e n , B r o n x , N Y 1 0 4 5 8 - 5 1 2 5 , U.S.A. P l e a s e i n q u i r e as to p r i c e s .
The Botanical Review 66(1): 1-56, January-March 2000 9 2000 The New York Botanical Garden 1
2 THE BOTANICAL REVIEW VI. VII. 2. Apocynoideae Burnett .. . . 38 a. Wrightieae G. Don . . . 39 b. Malouetieae Miill.-Arg . . . 39 e. Apocyneae Rchb .. . . - .. . . 40 d. Mesechiteae Miers . . . 41 e. Echiteae Bartl .. . . 41 3. Periplocoideae R. Br. ex Endl .. . . 42 4. Secamonoideae Endl .. . . 44 5. Asclepiadoideae R. Br. ex Burnett . . . 45 a. Marsdenieae Benth .. . . 46 b. Ceropegieae Orb . . . 47 c. Asclepiadeae (R. Br.) Duby .. . . 48 Acknowledgments . . . 51 Literature Cited . . . 52 I. Abstract
The Asclepiadaceae, as traditionally defined, have repeatedly been shown to be an apo- morphic derivative of the Apocynaceae. It has often been recommended that the Asclepi- adaceae be subsumed within the Apocynaceae in order to make the latter monophyletic. To date, however, no comprehensive, unified classification has been established. Here we pro- vide a unified classification for the Apocynaceae, which consists of 424 genera distributed among five subfamilies: Rauvolfioideae, Apocynoideae, Periplocoideae, Secamonoideae, and Asclepiadoideae. Keys to the subfamilies and tribes are provided, with lists of genera that (as far as we have been able to ascertain) are recognized in each tribe.
Zusammenfassung
Es wurde wiederholt festgestellt, dass die Asclepiadaceae in der traditionellen Um- grenzung ein apomorphes Derivat der Apocynaceae sind. Deshalb wurde oft vorgeschlagen, sie bei den Apocynaceae unterzubringen, damit die Apocynaceae eine monophyletische Gruppe darstellen. Bis jetzt wurde aber keine eingehende kombinierte Klassifikation pub- liziert. Hier stellen wir eine kombinierte Klassifikation vor for die Apocynaceae, mit 424 Gat- tungen, die in fiinf Unterfamilien gegliedert sind: Rauvolfioideae, Apocynoideae, Periplocoideae, Secamonoideae und Asclepiadoideae. Schliissel zu den Unterfamilien sowie den Triben werden gegeben. Die Gattungen, die zu jeder Tribus gehSren (soweit bekannt) werden aufgelistet.
II. Introduction
In 1810 Robert Brown published two seminal papers, "Prodromus Flora Novae Hollan- diae" (Brown, 1810a) and "On the Asclepiadeae" (Brown, 1810b). In these two publications Brown separated the Asclepiadeae (Asclepiadaceae) from the Apocineae (Apocynaceae) of Jussieu (1789) for the first time. In his Prodromus, which came out about a week before the second paper (Mabberley, 1985; Forster, 1991), he listed the taxa in two separate orders (families), which he called the Asclepiadeae and the Apocineae. His reasons for these changes were discussed in detail in the second paper: the main one being the presence of translators in the Asclepiadeae and their absence in the Apocyneae. Although Brown's classi- fication has been universally accepted and implemented, controversy over the delimitation of
CLASSIFICATION OF APOCYNACEAE S.L. 3
the two families has never been put to rest. They are clearly more similar to each other than to the rest of the Gentianales, and in a number of characters there is a gradation from the Apocy- naceae to the Asclepiadaceae. Three suggestions for recognition and expression of this rela- tionship have appeared: as one family (Hallier, 1905; Demeter, 1922; Safwat, 1962; Stebbins,
1974; Stevens, 1976; Thome, 1976, 1992; Judd et al., 1994; Struwe et al., 1994; Takhtajan, 1997); as an order separate from the Gentianales (Tsiang, 1934; Hutchinson, 1973); and as a suborder within the Gentianales (Rosatti, 1989; Nicholas & Baijnath, 1994; Omlor, 1998).
None of these possibilities seems to have met with much acceptance, so that, more than 180 years after Brown's (1810b) treatise, the group is still usually maintained as two families in the Gentianales. Nevertheless, new evidence from more detailed and extensive morpho- logical studies, as well as the rapidly growing body of molecular information (Judd et al., 1994; Endress & Albert, 1995; Sennblad & Bremer, 1996; Sennblad, 1997; Civeyrel et al., 1998; Sennblad et al., 1998; Potgieter, 1999), suggests that Brown's delimitation does not re- flect natural relationships. This new information and cladistic interpretations of it support the recognition of a single entity.
Here we again assess the relationship between the Apocynaceae and the Asclepiadaceae, and we present our arguments for the recognition of a single family. A new classification is proposed, descriptions of taxa down to tribal level are provided, and the genera at present rec- ognized in each tribe are listed. The priority rule has made necessary some changes of famil- iar names for subfamilies and tribes. We have adopted the oldest known names according to J. Reveal's Web site for suprageneric names (http://www.inform.umd.edu/PBIO). Thus some familiar names must be discarded. The major changes include: Rauvolfioideae Kostel. rather than Plumerioideae K. Schum.; Vinceae Duby rather than Rauvolfieae Bartl.; and Ceropegieae Orb. rather than Stapelieae Decne. Our classification is based mostly on mor- phological evidence accumulated through our own studies. Changes in classification based on results from molecular studies have been incorporated only where cladistic support was strong, or if they were also corroborated by morphological evidence.
III. Discussion
A. INFRAFAMILIAL CLASSIFICATION OF THE APOCYNACEAE S.STR.
The infrafamilial classification of the Apocynaceae s.str, has not changed much in the last 50 years. Two subfamilies are usually recognized: the Plumerioideae (= Rauvolfioideae), and the Apocynoideae. Additional subfamilies, such as Tabemaemontanoideae Stapf (1902), Apocynoideae sensu Woodson (1930), Cerberoideae Pichon (1948b), Carissoideae Endlicher (1838), have been described. However, either their delimitation is more ambiguous than that of the Rauvolfioideae and Apocynoideae, or they are at variance with our data. Consequently, they are not recognized here. The Rauvolfioideae typically have the corolla lobes sinis- trorsely contorted in bud, the anthers are mostly unspecialized and free from the style head, and there is a broad array of fruit and seed types, although the seeds are almost always eco- mose. The Apocynoideae, in contrast, are characterized by having the corolla lobes dex- trorsely contorted in bud and anthers specialized with lignified guide rails and adnate to the style head, forming a gynostegium (Fig. 1); the fruit is almost always a dry follicle with co- mose seeds. With the exception of Woodson's (1930) backward hypothesis, the Rauvolfioi- deae have usually been considered to be basal and more heterogeneous than the Apocynoideae. This view is also supported in all molecular investigations of the group and is followed here.
4 THE BOTANICAL REVIEW
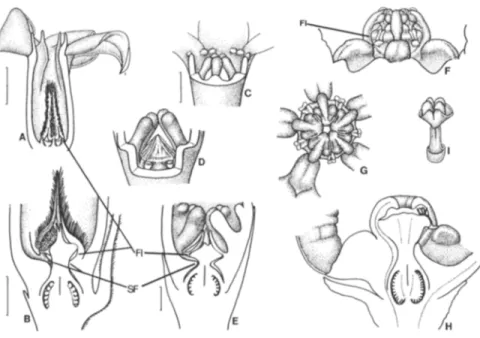
Fig. 1. Apocynoideae with filaments fused to corolla more or less at base, without any staminal foot:
Apocynum androsaemifolium (M. Endress). A. Flower. B. Dissected flower with two petals removed. C. Gynostegium. D. Base of dissected flower with one stamen removed. E. Half-flower (base only). F. Base of flower with stamen removed, showing small "corona lobe" on corolla (located below middle of each petal and pointing up into base of guide rail). FI = filament; N = nectary. Scales: A, B, 2 mm (at A); C, 1 mm (at A); D-F, 0.5 mm (at D).
The taxa of the Rauvolfioideae contain the least specialized flowers in the family. With the exception of the Tabernaemontaneae, which are characterized by the presence of lignified guide rails on the anthers, there are few distinguishing floral characters useful for differentiat- ing among tribes. The flowers are typically small and whitish and have a salveriform corolla tube. The anthers are small and ovate and are not adnate to the style head.
Allorge (1975) reported a retinacle in A llamanda and transferred the genus to the Echitoi-
deae (= Apocynoideae). However, the presence of a retinacle has not been confirmed in sub- sequent studies (Fallen, 1985), and a position in the Apocynoideae was not supported in more recent analyses either (Endress et al., 1996; Sennblad, 1997; Civeyrel et al., 1998). Leeuwen-
berg (1991, 1994) erroneously described Voacanga as having the anthers united with the style
head because the style and style head are shed with the corolla. More careful investigations re- veal that the style and style head come offwhen the corolla is shed not because the style head is united with the anthers but because of the conspicuously twisted corolla tube and the very large style head. On the inside of the corolla tube, the twisting in the tube manifests itself as tightly spiraled ridges that protrude into the corolla tube and reduce its diameter to much less than that of the enlarged style head. Thus, when the corolla is shed the ridges cannot pass over the style head and break it off instead. The only genus in the Rauvolfioideae in which the style
head is united with any other structure is Thevetia. In at least some species of this genus the
style head is postgenitally united with infrastaminal appendages of the corolla tube, below the anther. This union is not considered to be homologous to the retinacle and has been discussed elsewhere (Endress et al., 1996).
Thus, the very useful characters in the Apocynoideae provided by the structure of the retinacle are not found in the Rauvolfioideae. Obvious differences in floral structure charac- terize certain alliances but break down at tribal level. In constructing a tribal classification for
CLASSIFICATION OF APOCYNACEAE S.L. 5
the Rauvolfioideae, the systematist is faced with the dilemma of trying to find stable floral characters to distinguish the tribes amid a sea of monotonously similar flowers. For this rea- son tribal delimitation in the Rauvolfioideae has almost always been based on fruit characters. At first glance this seems like an excellent solution, because the fruits are distinctive and allow taxa to be easily categorized and keys constructed. Current knowledge, however, sug- gests that these fruit-based classifications are overly simplistic. As noted earlier (Fallen, 1983a, 1983b), it is always risky to place too much weight on a single character, especially when that character is more than usually subject to selective pressure. Recent studies (Endress et al., 1996; Sennblad & Bremer, 1996; Sennblad, 1997) indicate that fleshy, indehiscent fruits have evolved at least three times in the Rauvolfioideae. In addition, there are isolated cases of a taxon with fleshy, indehiscent fruits in a tribe that is otherwise characterized by de- hiscent fruits with a dry pericarp and winged seeds. (Examples include
Geissospermum
(fruit a berry) andVallesia
andMicroplumeria
(fruit a drupe)---all closely related toAspidosperma,
which has dry, woody follicles and flat seeds with a diaphanous wing (Potgieter & Albert, 1997, 1998a; Potgieter, 1999)--and
Melodinus
(fruit a berry), which is closely related toDiplorhynchus
and other, related, dry-fruited genera. This suggests a strong oscillatory or "back-and-forth" evolution between fleshy and dry, dehiscent and indehiscent fruits in re- sponse to dispersal by water, wind, or animals. Similar recurrences of certain characters in various angiosperm families has been reported by Armbruster (1996), Endress (1996), and Hufford (1997). The fruit characters should continue to be used, but with caution and only in conjunction with other characters. These fruit characters have been used so long that systema- tists have become lazy and do not search for new, meaningful characters. This is precisely what is now needed. In this paper we attempt to remedy this by delimiting the tribes of the Rauvolfioideae to reflect more accurately their true relationships. However, a consequence of this is that they are no longer as neatly and precisely defined as they were in older classifica- tions.The most comprehensive survey of the Rauvolfioideae was that of Pichon (1948a, 1948b, 1949, 1950b). It contained much new and detailed information and was a significant improve- ment over the classification published by Schumann (1895). However, tribes were still mostly based on a single fruit character. Pichon's ideas still dominate in floras and classifications to- day. For example, the most recent classification of Leeuwenberg (1994) scarcely differs from that of Pichon. Because they have been used for so long and the names are familiar, the tribes as recognized by Pichon and Leeuwenberg (cited above) will usually be outlined first (the names in quotation marks) in the discussion below, followed by the changes presented in the classification proposed here.
Pichon (1948a, 1948b, 1949, 1950b) split the rauvolfioid tribes into two main groups, de- pending on whether the inner pericarp was fleshy or dry. The first group contained the "Carisseae," "Ambelanieae" and "Macoubeeae" (fruit a fleshy, indehiscent berry) and the "Chilocarpeae" and "Tabernaemontaneae" (fruit consisting of dehiscent follicles with aril- late seeds). The second group contained the "Alstonieae" (= "Plumerieae" sensu Leeuwen- berg, 1994) (fruit a pair of dehiscent follicles with dry pericarp and usually winged seeds), "Rauvolfieae" (= "Alyxieae" sensu Leeuwenberg, 1994) (fruit an indehiscent drupe with a stony endocarp), and the "Allamandeae" (fruit a spiny unilocular capsule).
Syncarpy has generally been considered the plesiomorphic condition in the family, and the "Carisseae" was thus considered the most basal tribe. However, recent molecular results (Civeyrel, 1996; Endress et al., 1996; Sennblad & Bremer, 1996; Sennblad, 1997; Civeyrel et al., 1998) suggest that the traditionally defined "Carisseae" is not a natural group and that syn- carpy is perhaps not the basal condition in the family after all (Endress et al., 1996; Potgieter,
6 THE BOTANICAL REVIEW
1999). Rather, it appears that syncarpy has arisen independently in various assemblages and is, thus, not a particularly reliable character upon which to base a tribe. In the present treat- ment, the traditional "Carisseae" is split into three tribes.
Carissa
andAcokanthera
form the new Carisseae. Morphologically they are distinguished by having placentas that become in- durated in the fruit, forming a woody partition, as opposed to the remainder of the "Caris- seae," in which the placentas are unspecialized or pulpy in the fruit. The "Pleiocarpinae," characterized by apocarpous ovaries and conspicuous gaps in the corolla tube just above in- sertion of the stamens, is removed from the "Carisseae" and is treated as a separate tribe--the Hunterieae---as was proposed earlier (Fallen, 1986, as Pleiocarpeae). The remainder of the "Carisseae" (excludingMelodinus)
is, for the time being, maintained together as the tribe Willughbeeae. The monotypic "Chilocarpeae" is abandoned andChilocarpus
included in a newly defined Alyxieae.The other two tribes with indehiscent fruits and a fleshy inner pericarp recognized by Pi- chon (1948a) and Leeuwenberg (1994), the "Ambelanieae" and the "Macoubeeae," differ from the "Carisseae" in having specialized anthers with winglike margins formed into hard, lignified guide rails, such as are found in the Tabernaemontaneae as traditionally defined. Al- though the "Ambelanieae" and "Macoubeeae" have a syncarpous ovary and seeds embedded in pulp, whereas in the Tabemaemontaneae the ovary is usually apocarpous and the seeds dis- played with (often brightly colored) arils, the close relationship of these three tribes is well supported by morphological, chemical, and molecular evidence (Fallen, 1986; Zhu et al., 1990; Endress et al., 1996; Sennblad & Bremer, 1996; Sennblad, 1997; Civeyrel et al., 1998). Therefore, all are included here in a more broadly circumscribed Tabernaemontaneae.
Of the tribes of the Rauvolfioideae recognized by Pichon (1949, 1950b) that have a dry in- ner pericarp, the "Alstonieae" (= Plumerieae sensu Leeuwenberg, 1994) appears to be the most heterogeneous (Endress et al., 1996; Sennblad & Bremer, 1996; Sennblad, 1997). It is characterized by a plesiomorphic flower and fruit morphology, but several taxa have never been thoroughly studied. Preliminary molecular results indicate that winged seeds probably have arisen in parallel a number of times (Endress et al., 1996; Sennblad & Bremer, 1996; Potgieter, 1999). Here, the traditional "Plumerieae" (sensu Leeuwenberg, 1994) are split into three tribes. The "Plumeriinae," the "Cerbereae" (= the "Cerberoideae" sensu Pichon, 1948b), and the "Allamandeae" are combined to form the new Plumerieae. This is in accor- dance with independent studies, which suggest that the "Plumeriinae" is more closely related to the "Cerbereae" and the "Allamandeae" than to the remainder of the "Plumerieae" (Cop- pen & Cobb, 1983; Fallen, 1985; Nilsson, 1986, 1990; Endress et al., 1996; Sennblad & Bre- mer, 1996; Sennblad, 1997; Civeyrel et al., 1998). The "Craspidosperminae" (excluding
Strempeliopsis
andPlectaneia)
is here combined withMelodinus
as the new Melodineae. De- limitation of this tribe is somewhat imprecise. There is no single autapomorphic character known to be constant for all taxa. Rather, for the time being, circumscription must be based on a suite of characters and tendencies. These tendencies include: corolline corona often present, anthers sometimes with conspicuous sterile apical (sometimes also basal) append- ages, ovary often syncarpous to varying degrees, and pollen sometimes porate and sometimes in tetrads. The "Alstoniinae" and most genera of the "Aspidosperminae" are combined to form the new Alstonieae. Like the Melodineae, the Alstonieae is plesiomorphic and diverse. Common traits found in this tribe are alternate or whorled leaves, conspicuous gaps in the co- rolla tube above the level of stamen insertion, and the lack of a corolline corona. Finally, the "Catharanthinae" is transferred from the "Plumerieae" to a newly defined Vinceae, based on seed morphology (Endress, unpubl, data) and DNA results (Sennblad & Bremer, 1996; Senn- blad, 1997).CLASSIFICATION OF APOCYNACEAE S.L. 7
A source of confusion is the gaps in the corolla tube mentioned above as often present in the Alstonieae. The first report of these gaps was by Woodson (1951:124-125), who used them as a criterion to separate
Aspidosperma
(in which he found them to be present) from other genera, such asMicroplumeria, Geissospermum
andDiplorhynchus,
in which he found them to be "quite absent." Markgraf (1974:105-106) described similar gaps in the Madagas- can endemicStephanostegia
and raised the question of whether this was an indication of close relationship. Leeuwenberg (1997: 96), inexplicably, used this character in a key to separateStephanostegia
(gaps present) fromAspidosperma
(gaps absent). The gaps are due to the in- complete fusion of the petal primordia above the stamens (Boke, 1948; Nishino, 1982; Senn- blad et al., 1998:1147). The resulting gaps are especially long and thus conspicuous inAspidosperma
andStephanostegia.
However, they are also present, though shorter and not easily visible except in serial section, in a number of other taxa, includingCraspidospermum,
Geissospermum, Haplophyton, Pleiocarpa
andPicralima.
The "Rauvolfieae" of Pichon (1949) (= "Alyxieae" sensu Leeuwenberg, 1994) has tradi- tionally been characterized by having drupaceous fruits with a stony endocarp. Here the tribe is divided into the Vinceae---which normally have a style head that is vertically differentiated into functional zones (Fallen, 1986), fleshy mesocarp, nonruminate endosperm, flat seeds without a deep hilar furrow, and colporate pollen--and the Alyxieae---which typically have a style head that is uniformly receptive and secretory, stringy or woody mesocarp, ruminate en- dosperm, cylindrical seeds with a deep hilar furrow, and porate pollen. Although the Vinceae and the Alyxieae are superficially similar, taxa of the Vinceae cluster toward the basal part of the family, and it shows some affinities to the Tabernaemontaneae, whereas the Alyxieae ap- pears to be closer to the Plumerieae and Carisseae.
The second subfamily, the Apocynoideae, has generally been considered to be more ho- mogeneous than the Rauvolfioideae, and tribal delimitation is correspondingly more difficult (Fallen, 1986; Leeuwenberg, 1994). The most significant contribution to classification of the Apocynoideae was made by Pichon (1950a), who recognized four tribes: "Parsonsieae," "Nerieae," "Ecdysanthereae," and "Ichnocarpeae." These tribes were based mainly on the manner in which the anthers were united with the style head, a character Pichon (1948c) termed the "retinacle." Because the retinacle is very delicate and is easily ruptured during dis- section, characters based on its detailed structure have not been used by most subsequent apo- cynologists.
A number of revisions of Old World genera have been published by the Apocynaceae study group in Wageningen, under the leadership of A. J. M. Leeuwenberg (for references, see the literature cited in Leeuwenberg, 1983, 1988; Sennblad et al., 1998), and by Middleton (1994, 1995, 1997a, 1997b, and references therein). A detailed study by Endress et al. (1990) of the Holarrheninae (included in the "Plumerioideae" = Rauvolfioideae by Pichon, 1950b; De Kruif, 1981; and Leeuwenberg, 1983) resulted in the transfer of the three constituent gen- era
(Holarrhena, Carruthersia,
andSpirolobium)
to the Nerieae (sensu Leeuwenberg, 1988 = Wrightieae sensu Leeuwenberg, 1994) of the Apocynoideae. In the classification presented here all three genera are included in the Malouetieae. But broad, comparative studies of the group are lacking. In particular, the relationship of the Old World taxa with those in the New World has not been satisfactorily investigated, and this would be a rewarding focus for future studies of this subfamily.In his most recent classification Leeuwenberg (1994) stated, "The Apocynoideae genera are so closely interrelated, that it is probably impossible to place them into distinct tribes or subtribes." Nonetheless, in the same publication, he recognized three tribes: Echiteae, Wrightieae, and Apocyneae. Leeuwenberg's Echiteae and Wrightieae correspond for the
8 THE BOTANICAL REVIEW
most part to Pichon's "Parsonsieae" and "Nerieae," respectively; his Apocyneae is an amal- gamation of Pichon's "Ecdysanthereae" and "Ichnocarpeae."
Fortunately, interest in the Apocynoideae has revived in conjunction with the increasing use of molecular techniques. In a recent study of the Wrightieae, the three tribes of the Apocy- noideae recognized by Leeuwenberg (1994) were all shown to be paraphyletic, whereas the retinacle proved to be a useful character (Sennblad et al., 1998). In the present classification, five tribes are recognized in the Apocynoideae: Wrightieae, Malouetieae, Apocyneae, Echi- teae, and Mesechiteae; these are based mainly on retinacle structure and preliminary DNA re- sults and are augmented by other characters, as indicated in the key. The composition of the tribes differs significantly from those of both Pichon (1950a) and Leeuwenberg (1994). For example, genera that were previously included in the Echiteae by Leeuwenberg (1994) are here dispersed among three tribes. The relocations are, thus, too numerous to be discussed in- dividually. The five tribes of the Apocynoideae recognized here almost certainly represent a simplified scheme. The Apocyneae, especially, will probably need to be divided in some way, and some rearrangement of taxa will no doubt be necessary as more data accumulate. None- theless, until more adequate sampling is available for detailed studies, we feel this is the best representation.
B. RECOGNITION OF THE FAMILY PERIPLOCACEAE
After it was split from the Apocynaceae, the Asclepiadaceae was further divided into two families, the Asclepiadaceae and the Periplocaceae, on the suggestion of Schlechter (1905). This division was founded on the belief that in the Periplocaceae the pollen tetrads are loosely deposited on scooplike translators attached to a soft, sticky, somewhat amorphous viscidium, whereas in the remainder of the Asclepiadaceae the pollen is gathered into pollinia and at- tached to a cliplike corpuscle. The use of these two characters to separate these families was favored particularly by Bullock (1956), was supported by Hutchinson (1973) and Huber (1973), and has persisted to the present (see, for example, Kunze, 1993, 1996; Swarupanan- dan et al., 1996; Omlor, 1998). However, as the genera of the Periplocoideae have become better known, evidence has emerged that in many of them the pollen is gathered into pollinia (Verhoeven & Venter, 1998). We have personally seen this in
Decalepis, Gymnanthera,
andHemidesmus.
In addition, inDecalepis
the two pollinia produced in each locule of the anther tend to be pressed quite closely together, so that the appearance of a single polliniutlv--and hence only two on each translator--is created. This suggests the arrangement in the Asclepia- doideae, and the same pairing ofpollinia has been observed in three genera of the Secamonoi- deae:Genianthus
(Klackenberg, 1995b),Secamone
(Civeyrel, 1994), andSecamonopsis
(Civeyrel, 1996). In the Secamonoideae, the pollinia are similar to those seen in the Periplo- coideae, in that an outer wall is wanting. Therefore, the statement of Kunze (1996: 576) that "the jump from sticky tetrads to well-defined pollinia could be considered to be quite large," which was used as a justification for separating the Periplocoideae and the Asclepiadoideae at the level of family, is unjustified. Thus, in reality, the Periplocoideae and the remainder of the Asclepiadaceae are separated not by the presence/absence of pollinia but by the presence of a soft, sticky, more or less amorphous viscidium versus the presence of a harder corpuscle.It has also been said that the anthers are free from the style head in the Periplocoideae, whereas they are united to form a gynostegium in the Asclepiadaceae (Schumann, 1895; Meve & Liede, 1994; Swarupanandan et al., 1996). In all taxa of the Periplocoideae we have investigated, the anthers are clearly postgenitally fused to the style head (and this is shown here in Figures 2-7). Therefore, this claim is entirely without foundation. We can only sur-
CLASSIFICATION OF APOCYNACEAE S.L. 9
Fig. 2. Gymnanthera cunninghamii (P. I. Forster 6208): periplocoid with long corolla tube, staminal feet stretching down most of the tube but not thickening around neck of ovary. Pollen gathered into pol- linia. A. Bud. B. Flower with two petals removed. C. Gynostegium. D. Half-flower. E. Style removed to show staminal feet running down tube and alternating lower down with ridges of fine hairs. F. Pollinar- ium from above. G. Pollinarium from rear. fi = filament; sf= staminal foot. Scales: A, B, 2 mm (at A); C- E, 1 mm (at C); F, G, 0.5 mm (at F).
mise that the authors of these publications never actually looked closely at a periplocoid flower.
Another feature of the Periplocoideae is the lack of any lignified tissue in the margins of the anthers. The anthers are also not extended laterally toward the base into a swallow-tailed shape and therefore cannot form guide rails. This lack of guide rails is not matched in any other asclepiads except, to some extent, in Fockea: in Fockea the margins of the anthers are laterally closely adpressed to one another and are lignified toward the base, but they do not form a guide rail. This character is therefore somewhat equivocal for separating the Periplo- coideae from the Asclepiadaceae.
A staunch proponent of recognizing the Periplocaceae as a separate family is Kunze (1990, 1996). Much of his reasoning is based on his interpretation of the differences in the bases of the stamens between the Periplocoideae and the Asclepiadoideae. For the Periplocoi- deae he described the swollen region below the filaments as the "basal tube" and interpreted it as corolline or receptacular (Kunze 1990: 38). For the Asclepiadoideae he concluded that"the border between a receptacular base of the staminal column and an upper part in which the fila- ments are fused cannot be made out. In consequence the tubular part situated below the nec- taries is interpreted [as] receptacular in the whole family" (p. 44).
In his later paper Kunze (1996: 567) stated that the filament tube in the Asclepiadoideae has evolved by "replacement of the original free filaments by new elements originating from the inward[ly] protruding base[s] of the filaments." He stated as well that these results refute
10 THE BOTANICAL REVIEW
Fig. 3. Ischnolepis tuberosa (P. V. Bruyns 5963): periplocoid with more or less flat flower and corona
reduced to single lobes, staminal feet conspicuous but not forming tube around style. A. Flower. B. Gy- nostegium. C. Two anthers, viewed from inside, with staminal feet encircling their swollen bases. D. Half-flower. E. Pollinarium from above. F. Pollinarium from rear. fi = filament; sf = staminal foot. Scales: A, 4 mm; B, 1 mm; C, D, 1 mm (at C); E, F, 0.5 mm (at C).
Fig. 4. Cryptolepis grayi (P. Endress 9136): periplocoid with short corolla-tube, relatively long style,
very long filaments arising on short staminal foot, which forms a significant tube around the base of the style. A. Flower. B. Gynostegium. C. Half-flower. D. Pollinarium. fi = filament; sf = staminal foot. The staminal foot here begins to form a tube very similar to that in, e.g., Fig. 7E. Scales: A, 3 mm; B, 1 mm; C,
CLASSIFICATION OF APOCYNACEAE S.L. 11
Fig. 5. Raphionacmeprocumbens (P. V. Bruyns 4444): periplocoid with shorter corolla-tube, stami- nal feet beginning to enclose top of ovary. A. Flower. B. Gynostegium with one corona lobe cut away. C. Half-flower. 1). Pollinarium. FI = filament; SF = staminal foot. Scales: A, 2 ram; B, I mm; C, 0.5 mm; D, 0.25 rnm.
his hypothesis that the basal part of the "asclepiad filament tube is ofreceptacular origin" and that, consequently, the "basal tube in the Periplocaceae and the [basal part of the] filament- tube in the Asclepiadaceae are not homologous" (p. 573). This provided him with an addi- tional reason for recognizing the Periplocoideae as a separate family.
In the Periplocoideae the stamens each have a small, more or less cylindrical filament be- neath the anther. These filaments are inserted at different heights on the corolla tube but at- ways arise on the apex of a thickened ridge (which sometimes appears to be wrapped around the base of the filament and is usually deeply grooved below the filament). We shall term this ridge the "staminal foot," although we have found that, at the base of the stamen, it is impossi- ble to differentiate clearly between tissues derived from the stamen and those that are corol- line in origin. Nectar is secreted on the sides of this staminal foot, and in some cases the area between adjacent feet is also nectariferous. We have followed the development of this stami- nal foot, together with the length of the corolla tube, through several taxa in the Periplocoi- deae and have observed the following.
First, in species with a relatively long corolla tube, this staminal foot runs down the tube from the anthers and may fade out before the base (e.g., Gymnanthera [Fig. 2]; Raphionacme monteiroae, R. namibiana). It therefore does not necessarily have anything to do with the lowermost part of the tube and need not form any kind of"basal tube."
Second, in many periplocoids the corolla tube is much shorter, and in some the corolla is nearly flat (Ischnolepis [Fig. 3]). In cases where the corolla tube is shorter, the bases of these feet may become swollen around the neck of the style above the ovaries, initially just above
12 THE BOTANICAL REVIEW
Fig. 6. Periplocoideae: the formation of the staminal tube from the staminal foot. Ectadium virgatum
(P. V. Bruyns): periplocoid with separation ofcorolline corona at base of petals and staminal foot bearing the filaments, feet beginning to form staminal tube but still discrete. A. Dissected flower with two petals
removed. B. Half-flower showing base of tube. Cryptolepis oblongifolia (P. V. Bruyns): periplocoid with
separation of corolline corona at base of petals and staminal foot with filaments with conspicuous corona lobes at apex of each foot. C. Dissected flower with two petals removed. D. Further dissected flower with
one corona-lobe removed, showing anthers. E. Half-flower. Hemidesmus indicus (P. V. Bruyns):
periplocoid with staminal feet right up against style, completely enclosing it and ovaries and forming sta- minal tube; anthers fused to underside of style-head. F. Gynostegium from side. G. Gynostegium from above. H. Half-flower. I. Pollinarium. FI = filament; SF = staminal foot. Scales: A, 2 mm; B, l mm; C, 1 mm; D-H, 0.5 mm (at E), I, 0.25 mm (at C).
the ovary (Cryptolepis grayi [Fig. 4]; Raphionacme procumbens [Fig. 5]; Stomatostemma
monteiroae [Fig. 6]); then, with shorter style, they form a ring around the top of the ovary and
gradually fill up the space between the ovary and the style head (Cryptolepis oblongifolia, Ec-
tadium [Fig. 6]). The swollen bases that make up this ring are usually incompletely fused, so that the ring is undulate, with five mounds alternating with five slits (or at least epidermal sur- faces) between them.
Third, in extreme cases the limb of the foot disappears, and the filaments and anthers are pushed up against the style head. The lateral zone between adjacent feet fuses into a continu- ous ring around the style beneath the style head. Here all traces of epidermis between the swollen bases of the feet have disappeared, and a "staminal tube" around the style has arisen
(Hemidesmus indicus [Fig. 6]).
In our view, this arrangement leads directly to that found in the Secamonoideae/Asclepia- doideae, where the staminal tube has evolved further (Fig. 7D-G). In the Secamonoi- deae/Asclepiadoideae the anthers sit directly atop somewhat rectangular "filaments" (see Kunze, 1990: figs. 136-140; 1996: figs. 67-75). The partly to wholly fused inner, lateral mar- gins of these "filaments" form the so-called filament tube around the ovaries and style. These
CLASSIFICATION OF APOCYNACEAE S.L. 13
Fig. 7. Periplocoideae, Secamonoideae and Asclepiadoideae: staminal feet with filament, staminal tube without filament. Stomatostemma monteiroae (P. V. Bruyns): periplocoid with staminal feet near base of tube, corolline corona left behind at base of petals, ovary only partly enclosed by base of feet. A. Base of corolla tube with two petals removed. B. Half-flower. C. Pollinarium. Secamonopsis mada- gascariensis (P. K Bruyns): secamonoid with staminal tube on which anthers are sessile; outer corona encircling entire gynostegium. D. Dissected flower with two petals removed. E. Half-flower. Leptadenia reticulata (P. V. Bruyns): aselepiadoid with staminal tube on which anthers are sessile; flowers with co- rolline corona and outer corona encircling gynostegium. F. Half-flower. G. Dissected flower with two petals removed. FI = filament; SF = staminal foot; ST = staminal tube. Scales: A, 2 mm; B, G, 1 mm (at B); D, F, 0.5 mm (at B); C, E, 0.5 mm (at A).
filaments have a very unusual shape in comparison to those in the Periplocoideae (and the rest of the Apocynaceae). In view of the development of the staminal tube in some Periplocoideae and the nature of the filaments in this group, we find it altogether more plausible that in the Se- camonoideae/Asclepiadoideae the filament has been lost entirely and the so-called filament and filament tube are really a further development of the "staminal tube" that has appeared al- ready in some Periplocoideae. We therefore postulate that the Secamonoideae/Asclepiadoi- deae (Fig. 7) form a fourth stage in the cline outlined just above, namely:
Fourth, the staminal feet are attached to the corolla now only at a narrow base, and adja- cent feet are fused into a continuous ring around the style below the style head; that is, all traces of epidermis between their swollen bases have vanished, and they form a "staminal tube." The small, cylindrical filament is lost, and the anthers are consequently sessile on this staminal tube.
Another important fact is that while the situation described in the third stage leads natu- rally (in our view) to that in the Asclepiadoideae, the arrangement in first stage is similar to that in various Rauvolfioideae and Apocynoideae (cf. Fig. 8). In most Rauvolfioideae and
14 THE BOTANICAL REVIEW
Fig. 8. Apocynoideae with filaments fused to wall of corolla tube, with corolla tube thickened be-
neath them. A. Half-flower of young bud of Pachypodium succulentum (cultivated material).
B-C. Aganosma marginata (Fairchild Tropical Gardens). B. Part of flower with two petals excised and upper part of tube opened to expose anthers. C. Lower part of tube opened to show staminal feet and an- nular nectary around ovaries. In C a distinct staminal foot is present beneath the filament. FI = filament; N = neetary; SF = staminal foot. Scales: A, 1 mm; b, 2 mm; C, 1 mm (at B).
Apocynoideae there is also a small, more or less cylindrical filament beneath the anther. Moreover, there are also often ridges running down beneath each anther toward the base of the corolla tube (e.g., many Tabemaemontaneae, Apocynoideae).
Further aspects lend support to the existence ofa cline from the Periplocoideae to the Ascle- piadoideae. One is the position of the retinacle: When the filament is present, it lifts the anther above the thickest portion of the style head, so that the five anthers are connivent above it and are fused to the style head above its thickest portion (e.g., Fig. 4C and, especially, Fig. 5C). In
Hemidesmus, where the filament is particularly short, the anther is fused to the style head just beneath its thickest portion (Fig. 6H). This is typically the arrangement in the Asclepiadoideae.
The second aspect is the eorona. Kunze (1990) has also further differentiated between the Periplocaceae and the Apocynaceae on one hand and the Asclepiadaceae on the other by his conclusion that "the staminal corona in Aselepiadaceae is not homologous to the corona in
Periplocaceae and Apocynaceae" (1990: 45). The "corona lobe" in Raphionacme procum-
bens (see Fig. 5) clearly consists of three parts: a robust central lobule immediately behind the base of the filament inserted on the apex of the staminal foot, and two rather smaller lobules lateral to the central one. Kunze (1990) demonstrated clearly that these emergences are of co- rolline origin. However, in our view, a more important aspect is the fact that the robust, cen- tral lobule appears to be more intimately connected with the swollen staminal foot than the other two lobules and is more flexible in its position on the corolla tube than are the others.
In several other cases that we have examined (Decalepis, Cryptolepis, Ectadium, Stomato-
stemma [Figs. 6-7]), the apex of the staminal foot is some distance below the base of the pet-
CLASSIFICATION OF APOCYNACEAE S.L. 15
obscurely bifid (or bifid in its early stages), and there is no corona at the apex of the foot; in Cryptolepis this robust lobule arises more or less in the middle of the tube at the apex of the foot but some distance below the small pairs of corolline corona lobules. In Decalepis the ro- bust lobule is again a little away from the base of the petals, but here no lobules are left at the base of the petals. This situation again bears considerable similarity to many coronas found in the Asclepiadoideae.
The third aspect is the nectaries. In the Periplocoideae the nectaries are located on the sides of the staminal foot and/or in troughs between them; at any rate, below the filament. In the Asclepiadoideae the primary nectaries are located behind the guide rail at the top of the so- called filament tube and above the filaments. If this purported "filament tube" has evolved from the same staminal feet of the Periplocoideae, this would explain the presence of nectar- ies between the bases of the anthers and would remove the need to assume that nectaries have moved from below the filaments in the Periplocoideae to above them in the Asclepiadoideae.
Taking the above data into consideration, we fir'd (as suggested informally by Bruyns [ 1994]) that there is little morphological support for the separation of the Periplocoideae from the remainder of the asclepiads at the level of family.
Relatively little systematic work has been done on the Periplocoideae. Taxonomic revi- sions and pollen studies for a number of African genera have been published by Venter and Verhoeven (e.g., Verhoeven & Venter, 1988; Venter & Verhoeven, 1993, 1994; and refer- ences therein). In addition, four Malagasy genera were revised by Klackenberg (1997b, 1998,
1999). However, the Asian taxa remain especially poorly known.
Venter and Verhoeven (1997) published a much-needed comparative study of the Periplo- coideae, and for the first time tribes were delimited for the subfamily, based mainly on floral characters. Numerous superfluous genera were reduced to synonymy, and they recognized 44 genera (for a total of about 160 species), of which 17 are monotypic and a further 10 have just two species. We have incorporated into our classification the generic synonymy that Venter and Verhoeven propose.
However, for two reasons, we have not incorporated their tribes. First, although we are fa- miliar with relatively few of the species involved, those that do we know do not fit well into this classification. For example, in Decalepis nervosa the corolla tube is so short that the flower is more or less fiat; in Camptocarpus linearis the same is true. Yet both belong to genera in the Gymnanthereae, where the corolla tube is "well-defined and broadly campanulate to cy- lindrical" (Venter & Verhoeven, 1997:710). In addition, the flowers of Ischnolepis tuberosa are almost completely flat (Fig. 3), and we have been able to find almost no differences be- tween them and flowers ofPetopentia natalensis. Yet these two monotypic genera are placed in different tribes. Klackenberg (1999) also found that species of Petopentia fall into three dif- ferent tribes using the tribal classification of Venter and Verhoeven (1997) and thus doubted its value. Second, they conflict with the molecular results of Civeyrel (1996). There still seem to us to be far too many small genera. However, until detailed treatments of most of these taxa are available, it seems unlikely that any satisfactory subdivision of the subfamily can be achieved.
c. INFRAFAMILIAL CLASSIFICATION OF THE ASCLEPIADACEAE S.STR. 1. Recognition of the Secamonoideae
When Robert Brown (1810a, 1810b) separated the Asclepiadaceae from the Apocyna- ceae, he recognized three groups within the former: the "Asclepiadeae verae" (= Asclepiadoi- deae), the Periploceae, and an unnamed third group that contained only Secamone. Because it
16 THE BOTANICAL REVIEW
was unnamed, the status of this third group was ambiguous, and it was therefore subsumed in the Cynanchoideae (= Asclepiadoideae) in subsequent classifications by some authors (Ben- tham, 1876; Schumann, 1895). More commonly, however, it has been given equal rank (End- licher, 1838; Decaisne, 1844; Safwat, 1962; Bruyns & Forster, 1991; Liede & Albers, 1994). The Secamonoideae is a rather poorly known group. Nearly 80 species belong to
Secamone
(Goyder, 1992; Klackenberg, 1992a, 1992b), and there are at least 60 species inToxocarpus
and 16 inGenianthus
(Klackenberg, 1995b). There are also a few small genera (Klackenberg, 1995a, 1997a; Civeyrel, 1996).Morphologically, the Secamonoideae fall somewhere between the Periplocoideae and the Asclepiadoideae and share characters with both of these subfamilies. Although the flowers may occasionally be large (up to 100 mm in diameter in
Calyptranthera grandiflora),
they are mostly very small, and the constituent parts are similarly tiny. This applies especially to the minute pollinaria. As in the Asclepiadoideae, the pollinarium consists of a corpuscle and pol- linia. The corpuscle inPervillea, Secamone,
andSecamonopsis
arises as a single body that is U-shaped in cross section and grows vertically above the secretory fields. When fully devel- oped it is yellowish and somewhat waxy and soft (Omlor, 1996). This contrasts strongly with that in the higher asclepiads, where the corpuscle develops from two distinct parts, which later become fused across the base as the secretory fields themselves increase in size. In the higher asclepiads the corpuscle also has a rigid and much harder consistency.Although the pollinia were traditionally considered to be attached di[ectly to the corpuscle in the Secamonoideae, recent studies in
Secamone
(Civeyrel, 1995, 1996) have shown that in several cases there is a short caudicle between the pollinia and the corpuscle. Because the main distinction betweenSecamone
andSecamonopsis
was the absence/presence of a caudi- cle, this brings the distinctness of these genera somewhat into question.Furthermore, in the Secamonoideae each anther has four locules and consequently pro- duces four pollinia (as in the genera of the Periplocoideae that bear pollinia and unlike the Asclepiadoideae, in which each anther has only two locules). Civeyrel (1995, 1996) has made a number of interesting observations in the Secamonoideae, two of which suggest independ- ent trends toward reduction from four to two pollinia per pollinarium. In the first trend the pol- linia produced by one flank of an anther are closely adpressed to one another (as noted above for some Periplocoideae as well), thereby giving the impression that each anther produces only one pollinium on either side and that each corpuscle thus has only two pollinia attached to it. In the second trend, observed in
Secamone, Secamonopsis,
andTrichosandra,
one of the pollinia produced in each locule is much smaller than the other and is much more loosely at- tached to the corpuscle. In these cases, each pollinarium contains only two functionally effi- cient pollinia, which is reminiscent of the situation in the Asclepiadoideae. However, in the Secamonoideae with functionally bisporangiate anthers, it is the two inner (ventral) ones that are reduced (Civeyrel, 1995, 1996), whereas in the Asclepiadoideae it is the two outer (dorsal) ones that have become obsolete.A more detailed inspection of the pollinia of the Secamonoideae shows that they differ from those in the Asclepiadoideae in that they are composed of tetrads held together by cross- wall fusion, without an outer wall enclosing the pollinium. This is most similar to those genera of Periplocoideae with pollinia (Civeyrel, 1996; Verhoeven & Venter, 1998). It differs from the Asclepiadoideae, in which, in all members investigated to date, with the exception of Fockea (R. Verhoeven, pers. comm.), the pollinium comprises individual pollen grains and the whole structure is covered by a thick wall (the ectexine of Schill & J~ckel, 1978; Dannen- baum & Schill, 1991). Schill and J~ickel (1978) reported a thin outer wall enclosing the pollin- ium in
Secamone ligustrifolia,
but the composition of this wall was not studied in detail. ThusCLASSIFICATION OF APOCYNACEAE S.L. 17
it is uncertain whether it is composed of sporopollenin, like those in the Asclepiadoideae, or of elastoviscin, which Dannenbaum and Schill (1991) found covering the tetrads in Raphion- acme (Periplocoideae).
From the short discussion above, it is clear that the Secamonoideae are distinct from both the Periplocoideae and the Asclepiadoideae and deserve subfamilial status. It is by far the smallest subfamily, with only nine genera, all of which are included here in the single tribe Secamoneae.
2. Relationships within the Asclepiadoideae
In the Asclepiadoideae, four tribes are usually recognized: Marsdenieae, Stapelieae (here as Ceropegieae), Asclepiadeae, and Gonolobeae. Efforts have been made to refine the defini- tions of the tribes, especially by Woodson (1941) for the American taxa, Kunze (1995) for the Gonolobeae, Liede (1997) for the Asclepiadeae, and Omlor (1998) for the Marsdenieae. In the case of the Ceropegieae, which is probably the best known of these tribes, the difficulties were discussed at length in Bruyns and Forster (1991 ) and will not be repeated here. A listing of tribes and genera, which is nearly complete, was provided by Liede and Albers (1994).
One new tribe, the Fockeeae, has recently been separated from the Marsdenieae (Kunze et al., 1994). The Fockeeae was separated from the Marsdenieae by the lack of caudicles and the lack of a floor in the lower third of the corpuscle. Caudicles are lacking mostly in Genianthus and Secamone but may sometimes be present (Civeyrel, 1994; Klackenberg, 1995b), so that this seems to be a poor character on which to base a tribe. In addition, very frail corpuscles, which easily split longitudinally, are known in Eustegia, Gongronema, and Tylophora. In these genera it is the floor of the corpuscle that is weak, even though it may be present for most of the length of the corpuscle. In Eustegia (Bruyns, 1999) it has the same V-shaped profile that Kunze (1993:117) found in Fockea and observed as well in Tylophora. Thus this charac- ter, too, is not strong. Another character given by Kunze et al. (1994: 373), namely "stamens with large apical appendages," is true for Fockea but not for Cibirhiza, so it is certainly not definitive for the tribe.
A further argument that Kunze (1993) put forward to isolate Fockea from the remaining Asclepiadaceae was the hypothesis that the "lateral adhesive pads in Fockea" (attaching the corpuscle to the guide rails) are homologous to the "caudicles of other Asclepiadaceae" and therefore are unique to Fockea (and Cibirhiza). Kunze (1994: 232) has subsequently aban- doned this hypothesis and replaced it with the view that they are nonhomologous, which seems rather more plausible to us as well. In addition, Kunze (1990: 42-44) found that the co- rona in Fockea developed somewhat differently from that in other asclepiads. The ontogeny of the corona of Cibirhiza was never investigated, but it was claimed (Kunze et al., 1994: 373) that the basic construction of the corona is similar in both genera. Our own investigations have not confirmed Kunze's original conclusions about the differences in the development of the corona in Fockea and other Asclepiadaceae, and we have found that they follow more or less the same pattern (Fig. 9A-H),
In most features Kunze (1990, 1996) has demonstrated that Fockea occupies an interme- diate position between Secamone and the more advanced Asclepiadoideae. Similarly, it has pollinia made up of tetrads not covered by an outer envelope (i.e., resembling those in Seca- monoideae and those Periplocoideae with pollinia, rather than other Asclepiadoideae [R. Verhoeven, pers. comm.]).
An intermediate position for Fockea is also suggested by cladistic analyses of both mor- phological and molecular data in a study by Civeyrel (1996). In this study Fockea appears iso-
18 THE BOTANICAL REVIEW
Fig. 9. Coronal ontogeny in Fockea capensis (exhort. L. J. Hill). A. Very early stage with no corona
present (one anther knocked back in preparation). B. Beneath guide rail (GR, arrowed) a small swelling has begun to appear, which forms a "bay" beneath each guide rail with vertically descending parts be- neath each side o f the rail and a floor well below the rail; also a swelling beneath the anther but somewhat below it. C. The bay beneath the guide rail has increased greatly in size, and adjacent bays are connected by a low ridge; the inner corona (IC, arrowed) has begun to appear. D-F. Inner corona increases greatly in size; outer series differentiates into three different kinds of lobes: a broad one behind each inner lobe, two ascending ones rising up to touch the base o f the guide-rail (derived from the vertically descending parts in B) and an enclosing lobe below the rail (arising from the floor well below the rail in B). G--H. Structures that were clear in F begin to be obscured once more: anthers begin to disappear inside the tube formed by the corona, which is about half its mature size. GR = guide rail; IC = inner corona; OC = outer corona. Scales: all = 200/tm.
CLASSIFICATION OF APOCYNACEAE S.L. 19
lated at the base of the Asclepiadoideae. This could be interpreted as support for the tribe Fockeeae. However, this seemingly isolated position could also be due to inadequate sam- piing, for only two other species from among the 570 or so species of the diverse tribe Mars- denieae were included. In order to assess accurately the Fockeeae it would be necessary to include more putatively basal Marsdenieae. The tribe Foekeeae is therefore abandoned, with
Fockea
andCibirhiza
placed once more in the Marsdenieae.The most basal tribe of the Asclepiadoideae is the Marsdenieae. The Marsdenieae are mainly separated from the Ceropegieae by the absence of a hyaline insertion crest on the up- per or outer edge of the pollinium (if present, it lies on the inner or lower edge of the pollin- ium), and the more or less complete absence of an outer corona (Bruyns & Forster, 1991 ; Omlor, 1998). Together, the Marsdenieae and Ceropegieae are essentially separated from the Asclepiadeae by the orientation of the pollinia in the anthers: pendulous (below the corpuscle) in the Asclepiadeae; erect (above the corpuscle) in the Marsdenieae and Ceropegieae. The lat- ter is considered to be the primitive condition (Kunze, 1993: 120). Other characters that can be useful in deciding whether a genus belongs to the Asclepiadeae rather than to the Marsde- nieae or Ceropegieae are the presence of a narrow neck between the ovaries and the style head and of a constriction at the base of the sterile apical appendage of the anther, usually located opposite the corpuscles.
However, these characters do not exclusively separate these tribes. Some Ceropegieae (e.g.,
Orbea
andMacropetalum)
also possess a narrow neck between the ovaries and the style head, whereas some Asclepiadeae lack it entirely. In several Asclepiadeae the constriction at the base of the apical appendage of the anther is absent in mature flowers (e.g.,Astephanus,
Eustegia
andMicroloma),
although in most cases investigated it is present in early develop- mental stages of the flower. In most genera this distinction is clear, and they may be placed easily in one of these tribes. In some instances, however, this distinction is blurred.Some support for uniting the Ceropegieae and the Marsdenieae into a single tribe may be considered to be received from the molecular analysis by Sennblad and Bremer (1996). How- ever, in their study, out of the more than 500 species of Ceropegieae, two were sampled, and of the Marsdenieae (about 570 species), three species were tested. It is difficult to see how levels of sampling as low as this can possibly be considered to represent the position in such species-rich and diverse tribes, and this is not considered to constitute serious evidence that the two tribes should be united. Once further data have been amassed, the position should be reviewed.
In the case of
Tylophora,
N. E. Brown (1907-1909: 765) observed some species with erect and others with pendulous pollinia, and he confessed to being unable to decide which orienta- tion the pollinia had in yet other cases (as a consequence, he abandonedTylophoropsis,
which we follow below). With respect to its pollinaria,Tylophora
therefore is in limbo between the two tribes. InTylophora
the pollinia are mostly shortly ellipsoidal to nearly spherical, the cau- dicles are very thin and often short, and the corpuscle is mostly very small and frail. All of these suggest that this pollinarium is unspecialized and primitive and that it is from such structures that the more advanced pollinaria in the Asclepiadeae and Marsdenieae have arisen. In the species ofTylophora
that we have examined, we found a relatively long neck (short in a few cases) between the style head and the ovaries. In many there is also a horizontal incision between the anther and its apical appendage; it must be noted that this has not always been found in mature flowers (and perhaps needs to be searched for in small buds) and that it is more prominent in species with larger and more obviously pendulous pollinia. Additional characters listed by Kunze (1996) that support a position in the Asclepiadeae forTylophora
are: the lack of endothecial tissue in the guide rails, and the degree of reduction of the endo-20 THE BOTANICAL REVIEW
thecium around the locules and in the structure of the guide rails. Thus, despite the somewhat equivocal nature of these characters for
Tylophora
and its somewhat intermediate position be- tween the Marsdenieae and the Asclepiadeae, it is included here among the Asclepiadeae.Another awkward case is
Karimbolea verruculosa.
Here the shortly ellipsoidal pollinia are erect. The corona contains elements beneath the guide rails (an outer corona) and exten- sive inner parts and seems to be fairly typical of those found inCynanchum.
Unfortunately, none of the published illustrations (Liede et al., 1993) shows the relative position of these co- ronas on the staminal tube; nor do they show the style head and ovaries or the anther append- ages, which might assist in deciding on the systematic positionof Karimbolea.
Nevertheless, it seems that this species is merely an aberrant member ofCynanchum
(consequently belong- ing to the Asclepiadeae), which has retained the basal position of the pollinia. A position in the Asclepiadeae was also proposed by Liede et al. (1993) and accepted by Omlor (1998).Marsdenia
is by far the largest genus in the tribe (ca. 250 spp.) and also has the widest dis- tribution, being found in both the Old World and the New World. Various complexes of spe- cies have sometimes been treated as separate genera. Especially industrious in creating new genera were Schlechter (see Nicholas, 1992 for an enumeration of his many papers) and Bul- lock (1963). Taken over the whole range, however, many of the differences break down, and for this reason several of the segregate genera are mostly not recognized today. Both Rothe (1915) and Forster (1995) recognized a very broadly circumscribedMarsdenia,
and Forster's (1995) broad concept ofMarsdenia
is used here.The most comprehensive survey of the Marsdenieae is the recent doctoral dissertation by Omlor (1998), which provides an overview of the tribe. In addition to the characters men- tioned above that are commonly used to distinguish the Marsdenieae from the Asclepiadeae, characters of the seed coat were found to be useful (Omlor, 1998: 58-63). Of particular inter- est was the finding that the surface of the seed coat is smooth and hairless in the Marsdenieae. In contrast, the surface of the seed in "the
Tylophora
group" (includingTylophora, Merrillan-
thus, Pentastelma, Sphaerocodon,
andVincetoxicopsis)
has clusters of hairs of the type found in some genera in the Asclepiadeae. This is yet further support for the transfer ofTylophora
and these genera to the Asclepiadeae.A second group of genera that Omlor (1998) excludes from the Marsdenieae is what he re- fers to as "neotropical intermediate genera." This group includes
Anomotassa, Barjonia, Ble-
pharodon, .1obinia, Nephradenia, Petalostelma, and Vailia,
all of which he moved to theAsclepiadeae. W. D. Stevens (pers. comm.) is also convinced that most of these belong in the Asclepiadeae, so we have accepted this here.
The very large tribe Asclepiadeae is cosmopolitan but with particular concentrations of species in Africa and the New World. It consists of a group of genera surrounding
Asclepias,
which have large and conspicuously flattened pollinia (in fact the largest pollinia in the Ascle- piadoideae are found here inPachycarpus and Calotropis),
where pollinia are very obviously pendulous. The African component of this group was revised by N. E. Brown (1902-1903, 1907-1909), who united most of the genera underAsclepias.
It was partly revised by Bullock in the 1950s (for a listing of his relevant papers, see Dyer, 1975: 474), who tried to reorganize them into many smaller, segregate genera. Bullock gave up his attempt before it was com- plete. It was taken up again briefly by Kupicha (1984) but otherwise has languished in ne- glect, so the taxonomy of this part of the tribe is especially disorganized.The remaining, possibly more plesiomorphic genera, appear to form a cline from the larger pollinia of the
Asclepias
alliance toward the state inTylophora
and the other basal genera of the other tribes, where the pollinia are small and ellipsoidal, and it can be difficult to discern whether they are pendulous or erect. Liede (1997) has made a preliminary phylogenetic studyCLASSIFICATION OF APOCYNACEAE S.L. 21
of the Asclepiadeae, in which she defined the subtribes and reduced numerous genera to syn- onymy. However, Fishbein and Levin (1997) have indicated that there is little justification at this juncture to sweep so many of these little-known genera into synonymy until they have been more thoroughly studied. Most ofLiede's synonyms are not, therefore, incorporated into this classification.
The Gonolobeae are exceptionally poorly known, and even the recent survey by Vethacke (1994) fails to throw much light on how they may be separated from the other tribes. The char- acterization of this tribe in the key of Bruyns and Forster (1991) has been shown to be incor- rect by Kunze (1995). Thus it is necessary to fall back on the characterization by Woodson (1941), which is based entirely on features of the pollinarium: the horizontal (rarely pendu- lous) pollinia (which are broad rather than long), with a hyaline insertion crest and usually at least one partly concave face. Kunze (1995) has shown that only this insertion crest becomes lodged in the guide rail and that growth of the pollen tubes takes place through the concave face of the pollinium. We have verified this in Gonolobus gonocarpus, but it needs to be tested more widely across the tribe. Unfortunately, in many of the cases investigated, Veth- acke (1994) failed to show the natural position of the pollinia in the anthers, which, according to Woodson (1941), is one of the important features of the tribe. Another feature that Wood- son mentions is the sculptured faces of the pollinia; and this, too, is not very clear in many cases illustrated by Vethacke. In the few cases we have examined, and in the few instances in which one may make this out from Vethacke's illustrations, it appears that there is a horizon- tal groove at the base of the anther appendage and that the style head tapers off into the ovaries as it usually (but not always) does in the Asclepiadeae. Vethacke illustrated many of the folli- cles, and these appear also to resemble closely some of those in the Asclepiadeae. A case could, therefore, be made for subsuming the Gonolobeae within the Asclepiadeae. Swarupan- andan et al. (1996) proposed a division of the Asclepiadaeeae (which excluded the Periplo- caceae) into three tribes: Secamoneae, Stapelieae, and Asclepiadeae. The Marsdenieae were included in the Stapelieae; and the Gonolobeae, in the Asclepiadeae.
The Stapelieae were defined by pollinia attached to the caudicles at their bases, anther sacs not embedded in the tissue of the anther wings, anther wings always below the level of the an- ther sacs, and style head separated from the ovaries by a sharp constriction, with the gynoe- cium devoid of a "true style." In the Asclepiadeae, pollinia were said to be attached to the caudicles at their apex, anther sacs are partly embedded in the tissue of the anther wings, the anther wings form a collar around the anther sacs, and the gynoecium has "two true styles" and lacks a constriction beneath the style head (Swarupanandan et al., 1996: 362).
It is now fairly well known that the guide rails (anther wings) are basal elongations of the dorsal pollen sacs (Kunze, 1996; Omlor, 1996). Therefore, it is impossible for the anther sacs to be "embedded in the tissue of the anther-wings." We are also quite unable to establish any justification for the statement that the anther wings form a"collar" around the anther sacs, and
nowhere do the authors show what they mean by this "collar."
The statement that the pollinia are attached to the caudicle at their apex is not correct for the Asclepiadeae. The many pollinaria illustrated by Kupicha (1984) demonstrate clearly how variable the position of attachment of the caudicle to the pollinium may be in the Asclepia- deae: from the apex to near the middle of the outer edge. Kunze (1995) has shown that the pol- linia in the Gonolobeae are laterally elongated (i.e., broad rather than long) and are attached to the caudicle not at their apex but near the middle of the outer edge. In the Ceropegieae s.str. the caudicle is not attached at the base of the pollinium. This is often true in the Marsdenieae (e.g., in Stigmatorhynchus, Bruyns, 1995), but in the Ceropegieae the attachment is usually toward the base but below the outer edge (see also Bruyns, 1995 for examples).