The role of cell death and myofibrillar damage in contractile dysfunction of long-term cultured adult cardiomyocytes exposed to doxorubicin
Texte intégral
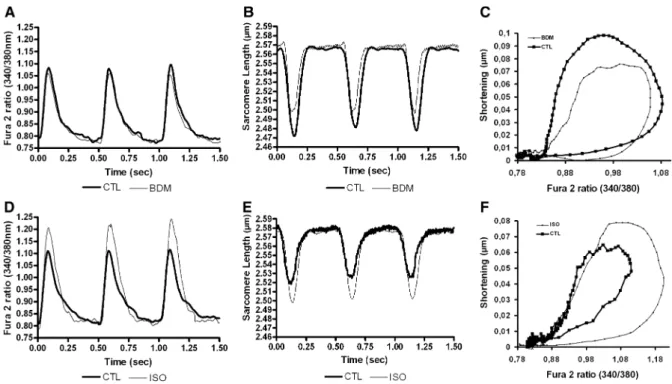
Figure


Documents relatifs
Theoretical SHG-IP (dotted lines) is obtained from the myofibrillar schematic view for a laser beam focused and displaced along the horizontal line at the center of the
In this paper, we choose to describe the typical behavior described above by means of a viscoelastic-damage constitutive model where the framework of the continuum damage mechanics
Consommation problématique d’opioïdes signifie aussi : • Utiliser un médicament opioïde de manière inappropriée, comme en prendre plus que prescrit ou au mauvais moment
Information Elaborée conjointement par l’équipe du Centre MITIC interjurassien et des enseignant-e-s des deux cantons, cette publication est destinée à mieux faire connaître
derbundskrieg 1847 ins Ausland, von wo er erst nach 30 Jahren wieder ins Wallis zurückkehrte", während der eifrige Saaser Chronist Peter Josef Im- seng (1796-1848) 1848 in
In the Falck case, the far-sighted family champion of change Alberto Falck—with crucial support of the external CEO Achille Colombo—was able to de-escalate the family business
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
l’utilisation d’un remède autre que le médicament, le mélange de miel et citron était le remède le plus utilisé, ce remède était efficace dans 75% des cas, le
