Task-dependent influence of genetic architecture and mating frequency on division of labour in social insect societies
10
0
0
Texte intégral
Figure
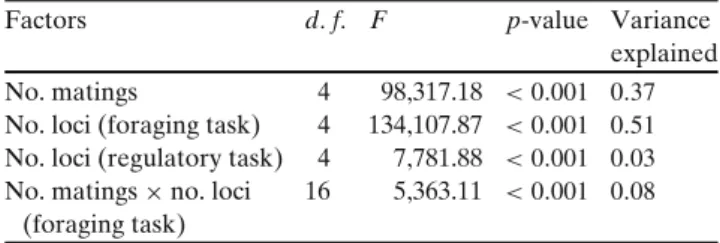

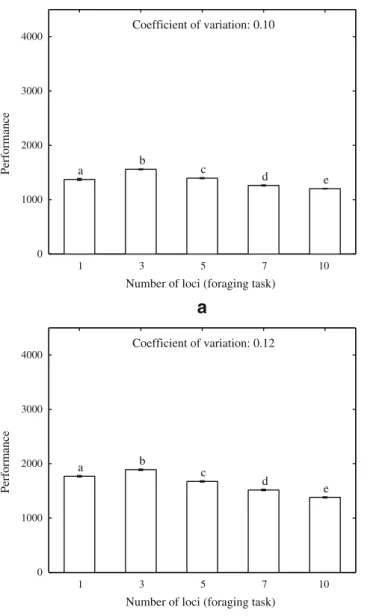
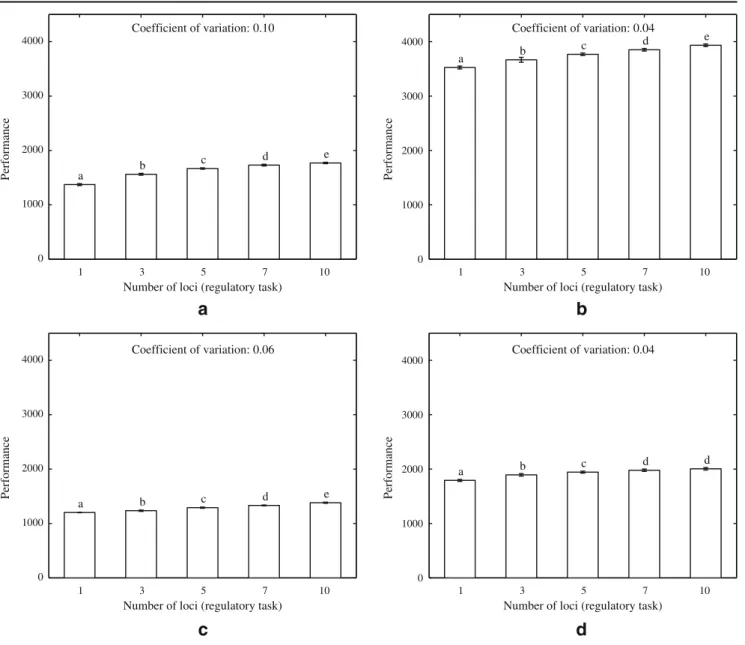
Documents relatifs