Effects of Long-Term Trampling on the Above-Ground Forest Vegetation and Soil Seed Bank at the Base of Limestone Cliffs
9
0
0
Texte intégral
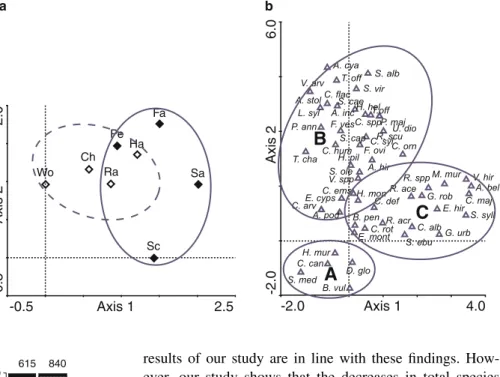
Figure

Documents relatifs