Design of a multifunctional biomineralized armor system : the shell of chitons
121
0
0
Texte intégral
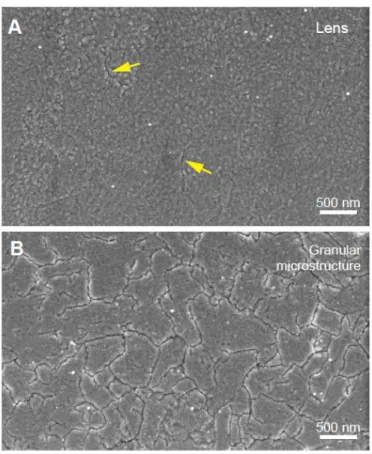
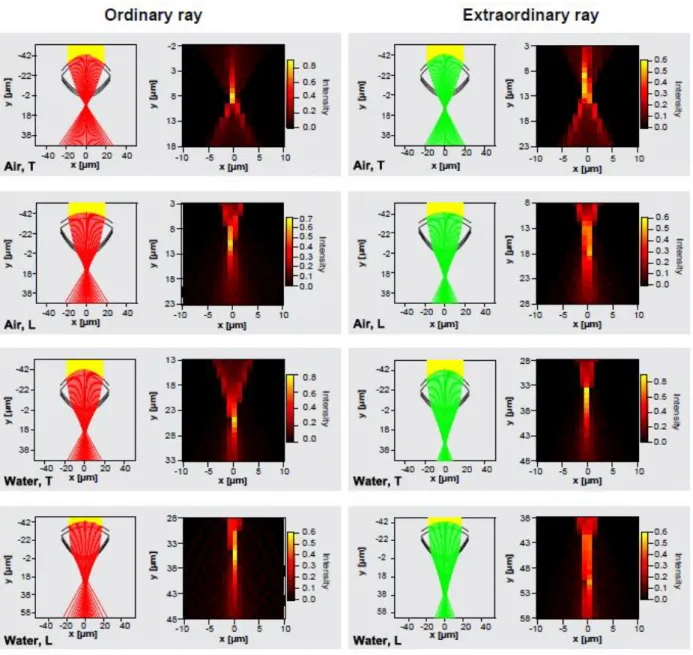
Figure

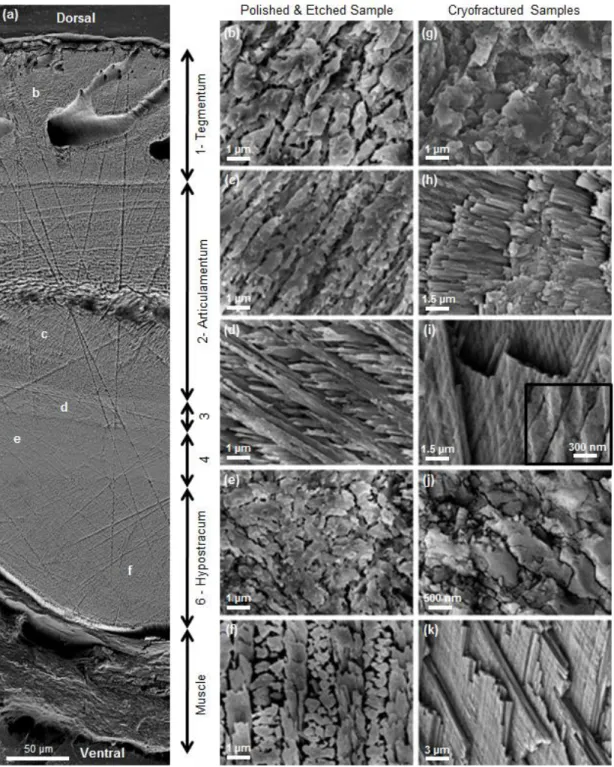
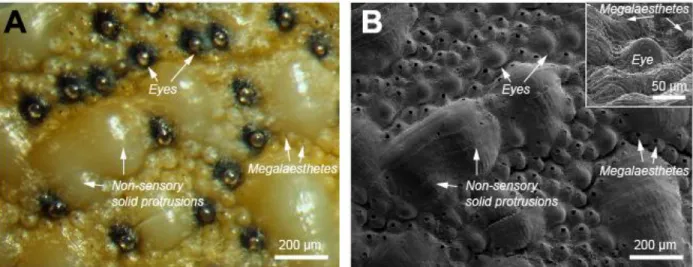
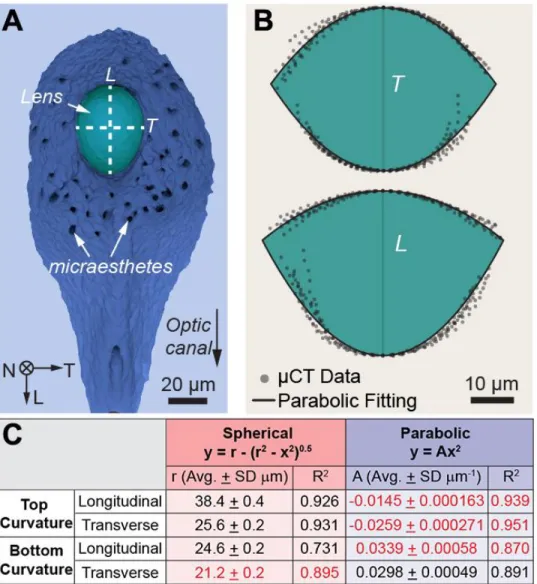
+7
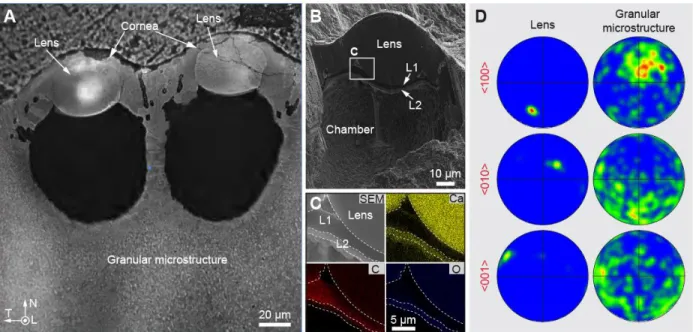



Documents relatifs