Mice deficient for the epidermal dermokine and isoforms display transient cornification defects
Texte intégral
Figure

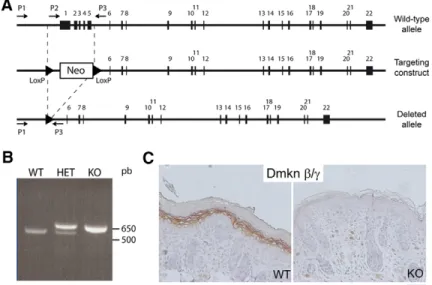

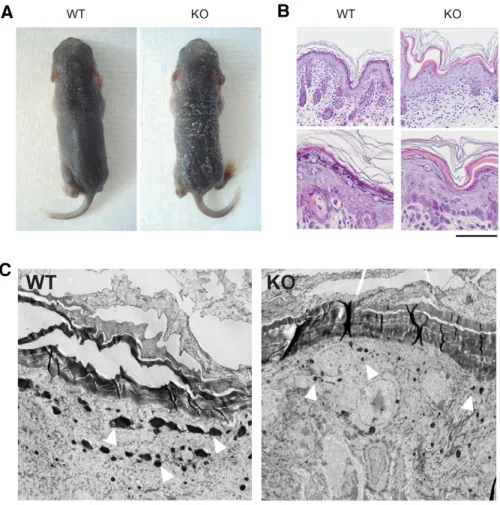


Documents relatifs
for this particular filtering characteristic, 16 of the 48 solutions are real-valued. Only the real solutions have a physical interpretation and are therefore
In a sys- tematic approach to find the best-suited ref- erence genes for qRT-PCR in honey bees, we analyzed the behavior of four candidate refer- ence genes (act, rp49, ef1-alpha
Residual risk (RU) associated with region-level proportion of samples positive to black queen cell virus (BQCV, a), to deformed wing virus (DWV, b), and to Kashmir bee virus (KBV, c)
After a brief overview of the impact of this polymorphism on different traits, we present thereafter preliminary studies, which rely upon a candidate gene approach followed by a
La MPR a développé une approche globale du patient, en s’appuyant notamment sur la Classification internationale du fonctionnement, du handicap et de la santé (CIF), qui
à l’école; les cloches et les messages intempestifs ont disparu, les activités complémentaires à l’enseignement sont concentrées dans des.. La recherche de la qualité:
The questions addressed in the present study were: (a) how many healthy calves born from seropositive mothers were precolostrally positive (thus indicating prenatal infection); (b)
The system is not useful for the retention force measure- ment of double crown retained dentures as a tilting of the denture could pretend excessive forces not clinically existing..
