Prolonged mechanical ventilation worsens sepsis-induced diaphragmatic dysfunction in the rat
Texte intégral
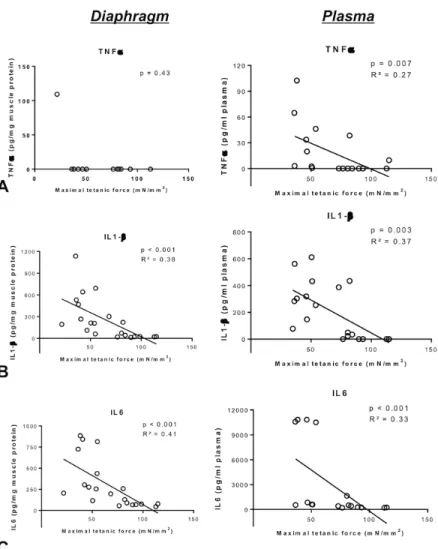
Figure

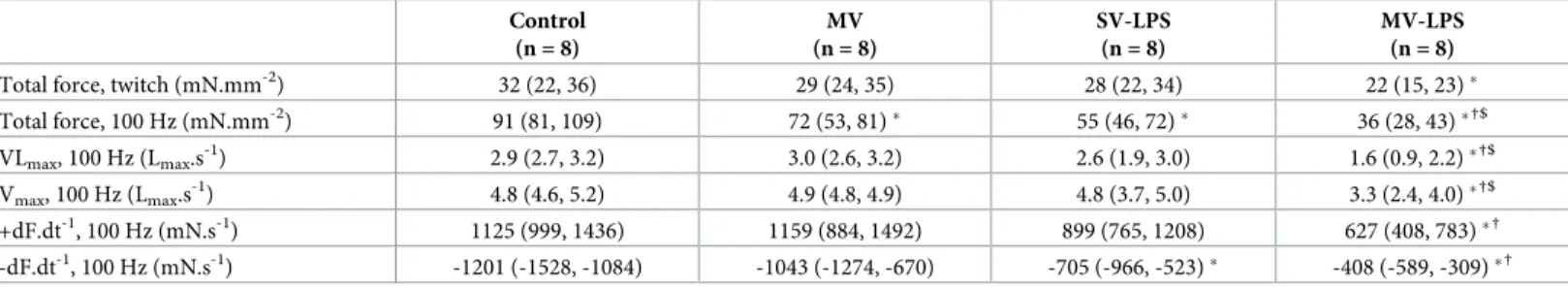
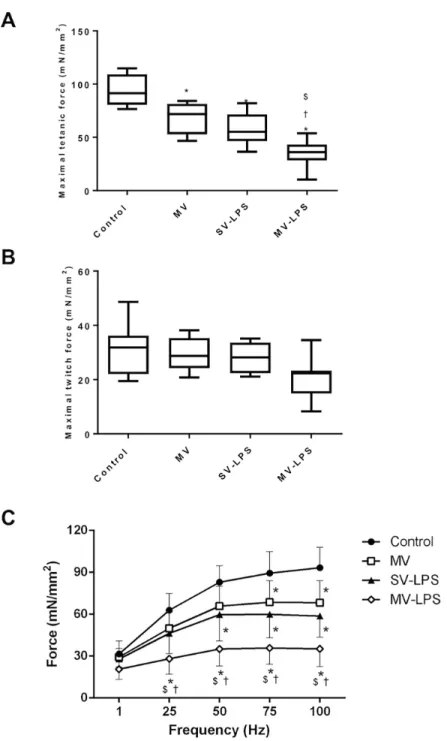
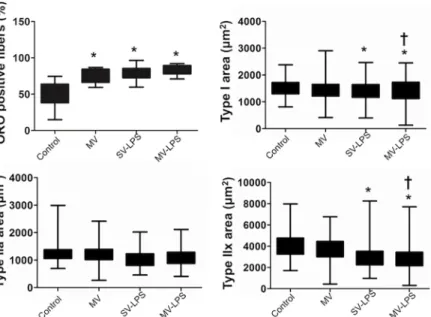
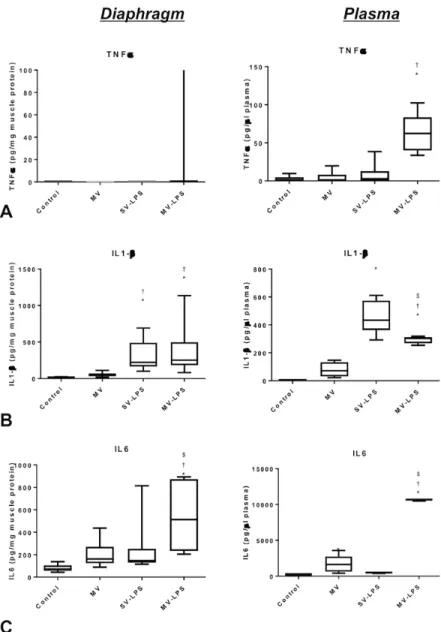
Documents relatifs
recognised as effectively as samples A and C in the assay of 5B in comparison with other assays indicating that antigenic determinants in the TNF - molecule are
Plusieurs stratégies peuvent être utilisées pour réduire le risque de réactions immédiates et retardées à la perfusion : traitement d’induction à 3 doses (0, 2 et 6
Le Panel sclérodermie, effectué à l’aide de la trousse EUROLINE MC Systemic Sclerosis (Nucleoli) Profile (IgG), a comme objectif la détection des autoanticorps (AAC) humains de
The design of WIPO Pearl seeks to offer users flexible and distinct yet complementary ways of search- ing the terminology dataset: a traditional search by term, called
Moreover, while Nazlioglu and Soytas (2012) focused on agricultural commodities with first generation panel unit root test which have the restrictive assumption to not
In this latter view, the important physics is a Mott type transition with lattice parameter involving not an occupation number change but rather a change in the
RT3: More longitudinal observational studies & more empirical works RT4: How to create an ecosystem RT5: Become more relevant to industry RT6: What can we learn from other
When a strong factor assumption is imposed and the number of factors is assumed to be fixed, the proposed two-step estimator is found to be asymptotically normal under weaker
