A Data Integration Multi-Omics Approach to Study Calorie Restriction-Induced Changes in Insulin Sensitivity
Texte intégral
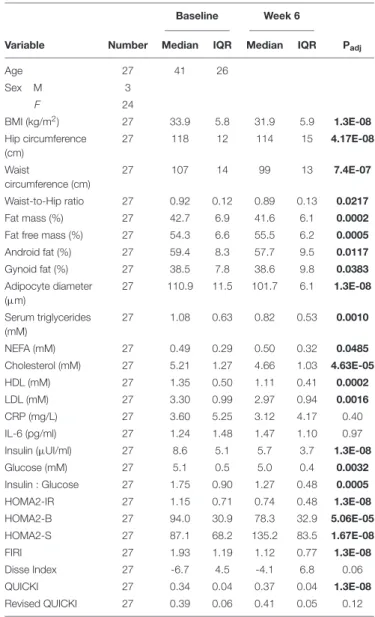
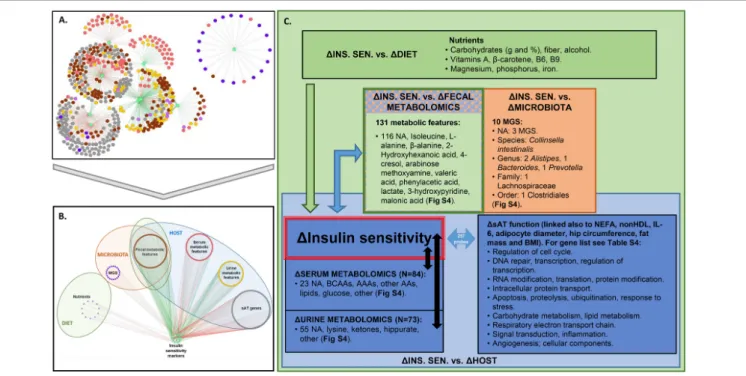
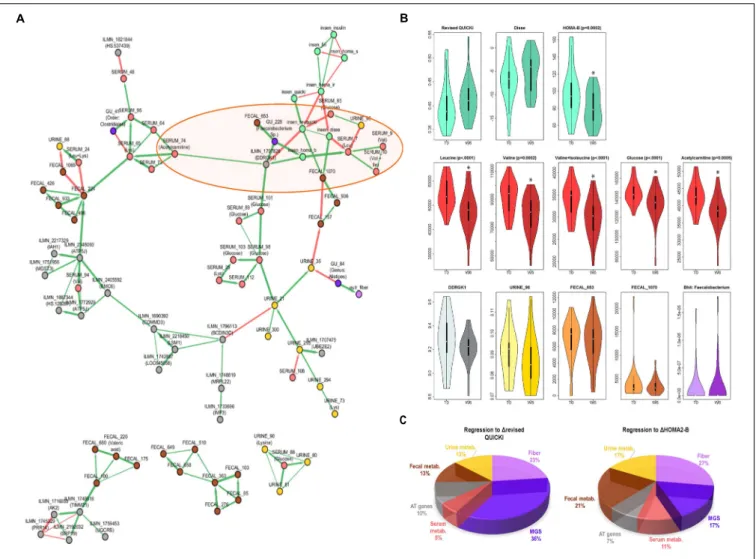
Figure

Documents relatifs
IRIS integrates two well-known public data sources MIRC and MyPacs and two medical ontologies RadLex 6 and The Systematized Nomenclature of Medicine Clinical Terms (SNOMED CT) 7.
Con- sequently, the documents serve as input for training vector representations, commonly termed as embed- dings, of data elements (e.g. entire relational tuples or
Additional file 12: Ovarian cancer multi-omics data visualisation on ACSN: zoomed on mitochondrial metabolism signalling pathway. Immunoreactive (A) and Proliferative (B) subtypes
We have shown that the query response time in this approach is better than that of fully virtual data integration approach and the data freshness is much better that of a
This Article should have been published under a Creative Commons licence according to the Nature policy on publishing the primary sequence of an organism’s genome for the first
Simultaneous analysis of distinct Omics data sets with integration of biological knowledge: Multiple Factor Analysis approach... BMC Genomics, BioMed Central, 2009,
Keywords: Multiple omics data integration, Multivariate factor analysis, Missing individuals, Multiple imputation, Hot-deck imputation..
Identification of causal signature using omics data integration and network reasoning-based analysis.. Méline Wery, Emmanuelle Becker, Franck Auge, Charles Bettembourg, Olivier
