Autocrine-controlled formation and function of tissue-like aggregates by hepatocytes in micropatterned hydrogel arrays
Texte intégral
Figure
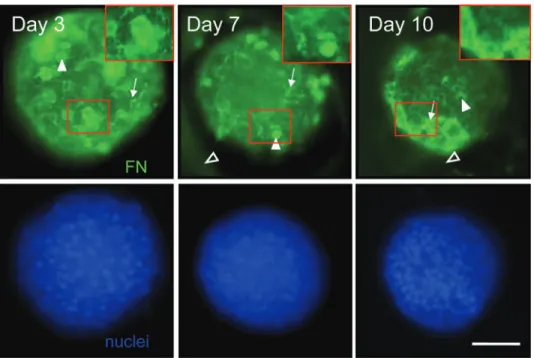
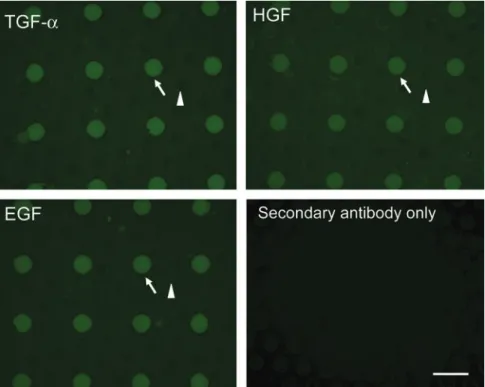
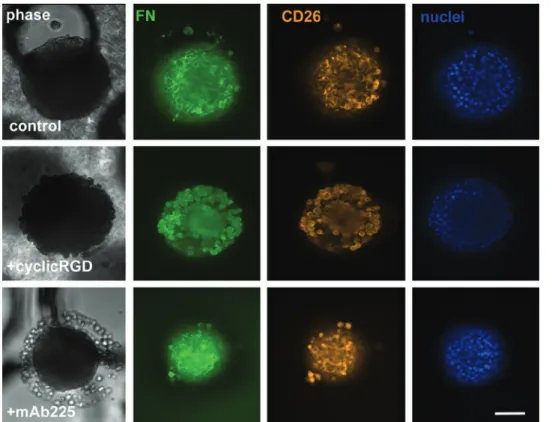
Documents relatifs
The global expression of main hepatic drug transporters at the mRNA level, associated to detectable activities of NTCP, OATPs and OCT1, was moreover observed in
In this paper, for the first time, ethanol was shown to act reciprocally with B[a]P in promoting hepatocyte oxidative stress and cell death through a novel mechanism of
Our study provides two new pieces of evidence indicating the precursor- product relationship between UDP-glucuronate and glucu- ronate since (a) two agents (resorcinol and
Hence, the aim of the present study was to analyze whether the following phase II resveratrol metabolites, trans-resveratrol-4 0 -O-glucuronide (R-4G),
Three antioxidant models namely DPPH radical scavenging activity, hydroxyl radical scavenging activity and metal ion chelating ability were used to reflect the
Devant ]a lenteur de la constitution de I'organisme de liquidation' le Conseil Municipal, lors de la séance du 25 Octobre 1946 (157)' rappeile la désastreuse situation financière
2)) L L''iin nd diiccaatteeu urr m meen nssu ueell d dee ll''eem mp pllo oii iin nttéérriim maaiirree een n ffiin n d dee m mo oiiss est élaboré par la Dares à partir
After incubation for 3 h with [11,12- 3 H] lauric acid (4 nmol and 3.7 MBq per dish), 2-D electropho- resis and western blotting followed by spot exci- sion, each protein was
