3′-End processing of pre-mRNA in eukaryotes
19
0
0
Texte intégral
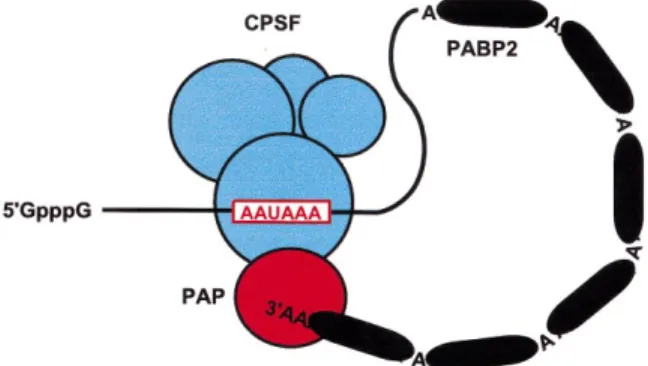
Figure
Documents relatifs
