Reward processing under influence : Effects of stress and cognitive load on reward processing, and their clinical implications for the vulnerability to major depression
Texte intégral
Figure
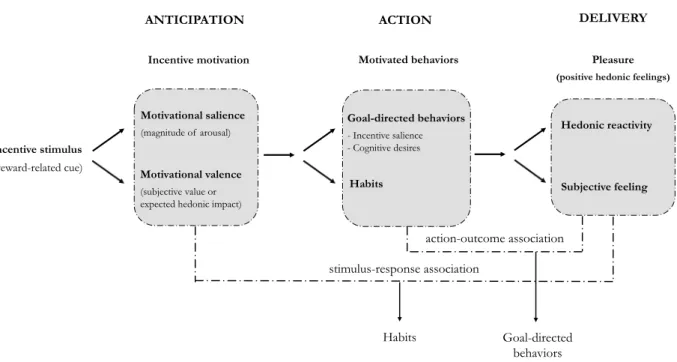
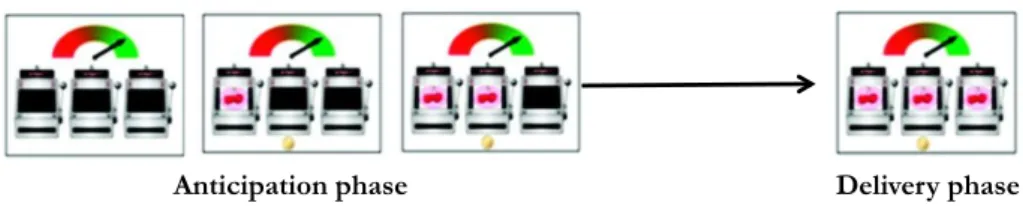
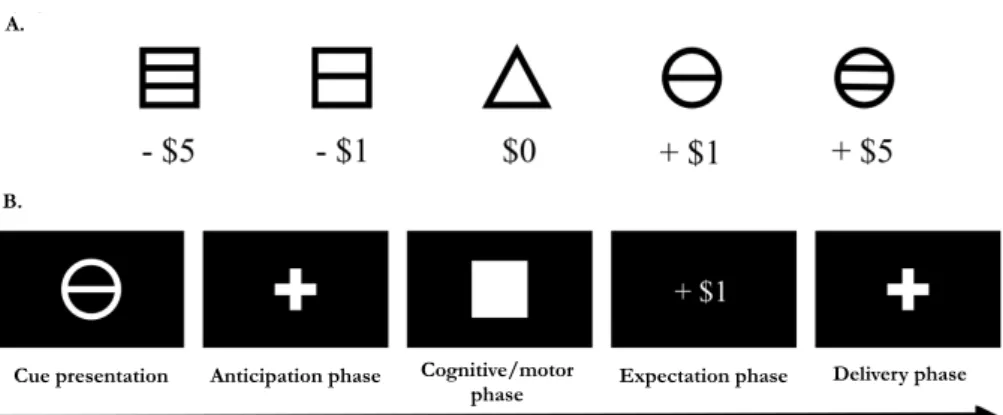
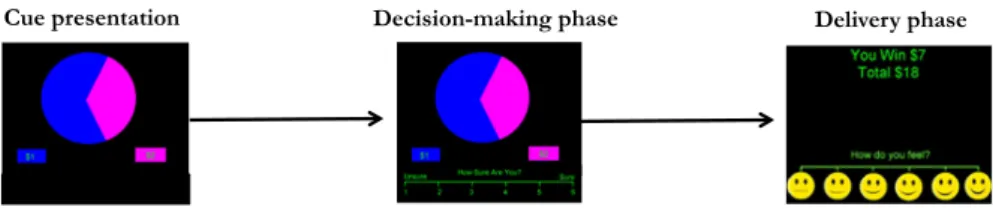
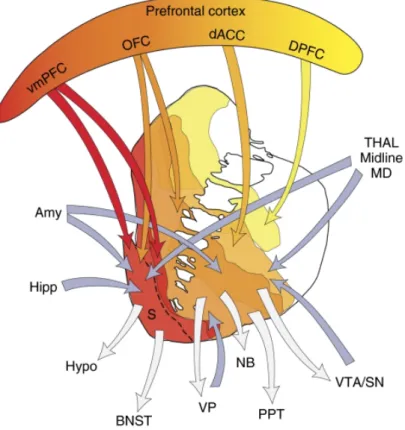

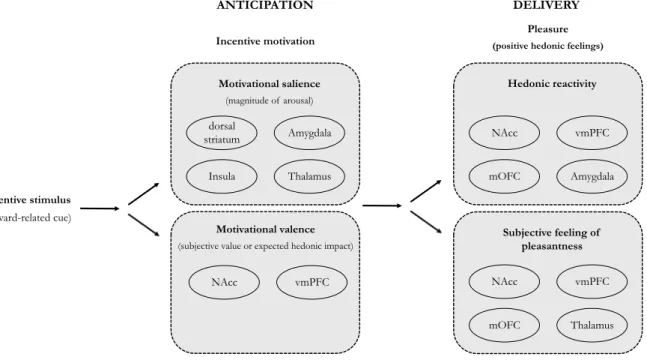

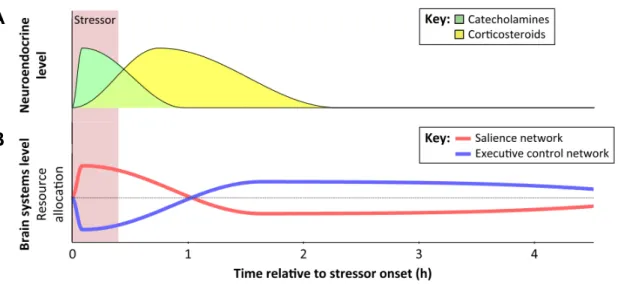
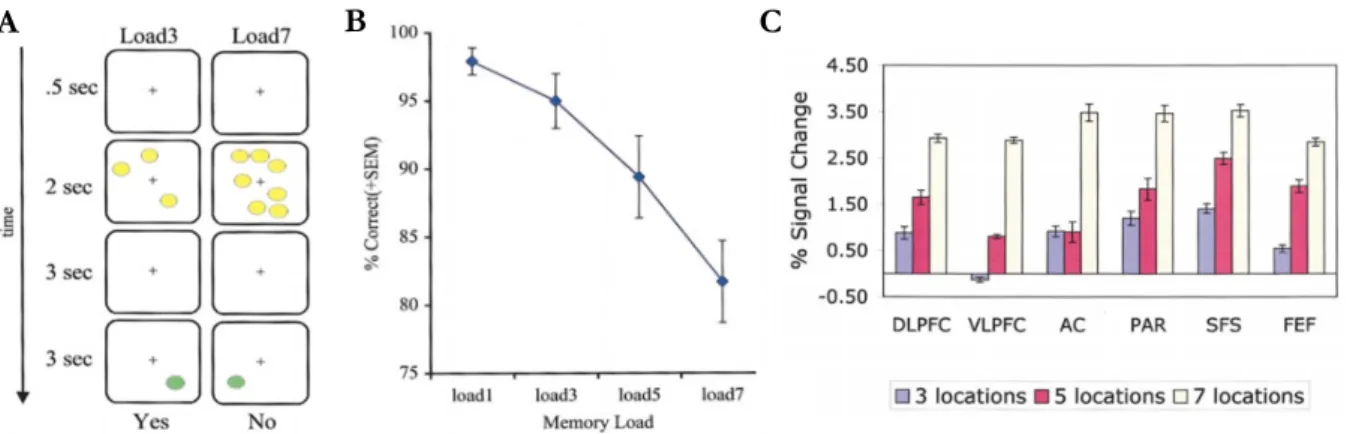
Documents relatifs
Null Hypothesis: There will be no difference in biomechanical or neuromuscular features (spine posture, muscle activity, perceived pain rating, spine flexibility and muscle
The greater P300 amplitudes observed when the participants are chosen by the proposer suggest an increase in attentional resources allocated to the task when the participants
Beha- vioral reaction times, reward-anticipating licking dura- tions and error rates varied for the same reward when tested in two different reward combinations, although the
mPF C: medial prefrontal cortex; CeA: Central Amygdala; BLA: Basolateral nucleus of the Amygdala; vSUB: ventral subiculum; BNST: Bed nucleus of the Stria Terminalis; dCA3:
An experiment for the study of the effects of exposure to a winter-like photoperiod on the diurnal rodent Arvicanthis ansorgei indicated that shortened day length with
First, undocu- mented migrants participating in our study may not be representative of the total undocumented population in Geneva because of the predominance of Latin American women
À travers une lecture qui se veut détachée de la seule figure de l’auteur comme principe d’ana- lyse et d’interprétation de l’écriture romanesque d’Abdellah Taïa,
REWARD can help improve interoper- ability and integration among different reward schemes employed by different service providers and define a uniform process for creating
