Identification of chlorinated solvents degradation zones in clay till by high resolution chemical, microbial and compound specific isotope analysis
14
0
0
Texte intégral
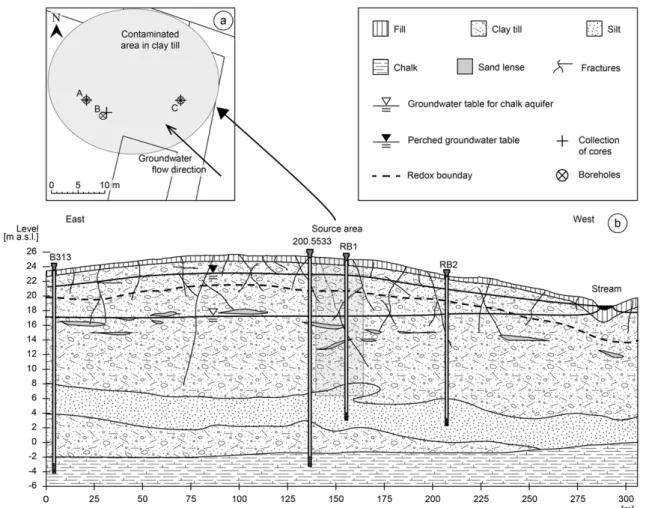
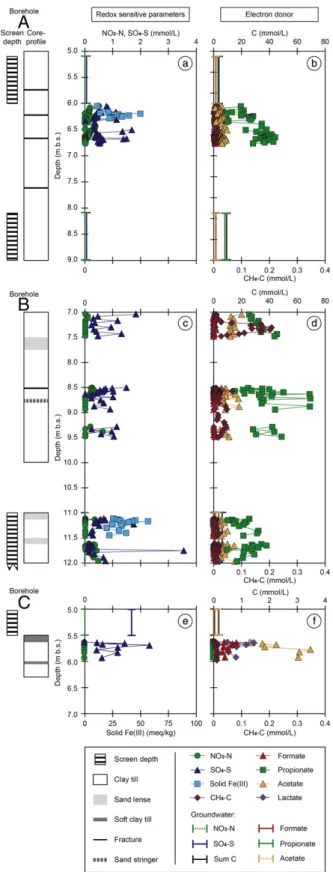
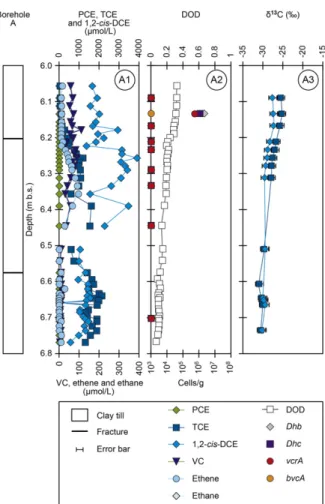
Figure
Documents relatifs