Detection of the ADP-ribosyltransferase toxin gene (cdtA) and its activity in Clostridium difficile isolates from Equidae
Texte intégral
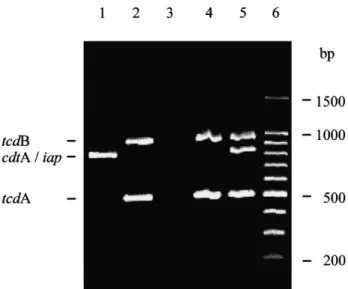
Figure
Documents relatifs
611 فيرعت نكمي ًاحلاطصاو ًةغل باهرلإا موهفم طبض دعبو باهرلإا يلودلا ىلع اذهو ،ًايلود ًاعباط وأ ًادعُب ذخأي يذلا باهرلإا كلذ ":هنأ :يف
In addition, to assess if females would re-mate with the same male, a third group of seven mated pairs were kept together in individual cages after their first mating
The fine-scale genetic structure of the malaria vectors Anopheles funestus and Anopheles gambiae (Diptera: Culicidae) in the north-eastern part of Tanzania... The fine-scale
To cite this version : Benammar, Bouchra and Thomas, Nathalie and Boucheret, Marie-Laure and Poulliat, Charly and Dervin, Mathieu Analytical Expressions of Power Spectral Density
L’accès à ce site Web et l’utilisation de son contenu sont assujettis aux conditions présentées dans le site LISEZ CES CONDITIONS ATTENTIVEMENT AVANT D’UTILISER CE SITE WEB.
L’accès à ce site Web et l’utilisation de son contenu sont assujettis aux conditions présentées dans le site LISEZ CES CONDITIONS ATTENTIVEMENT AVANT D’UTILISER CE SITE WEB.
Cette confrontation plus ou moins douloureuse avec les points de vue de sa famille, de l'institution scolaire ou professionnelle et selon les rapports sociaux, culturels et sexués
Emotions and reason are therefore on a par in the domain of moral enhancement, and if we accept improvement of the second through direct cognitive enhancement (like drugs), we
