Climate reconstruction from pollen and δ 13C records using inverse vegetation modeling – Implication for past and future climates
Texte intégral
Figure
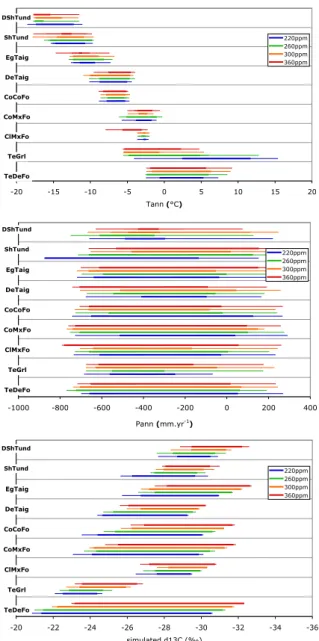
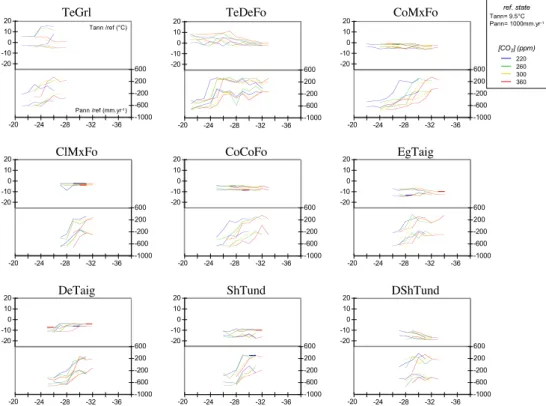
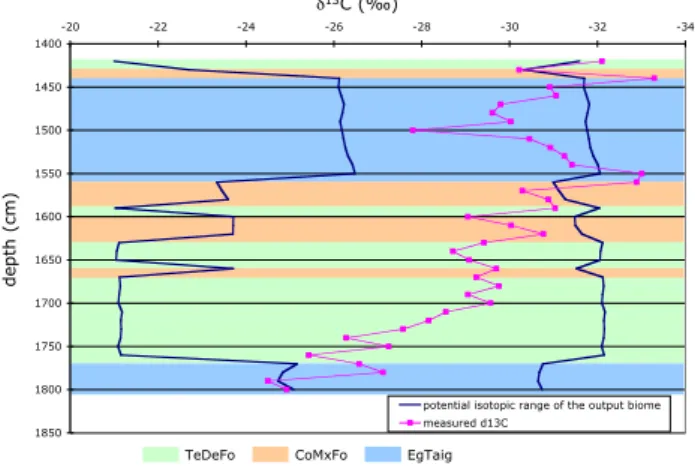
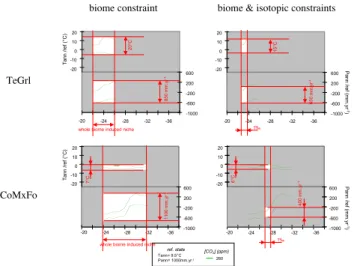
Documents relatifs
Fuzzy disaggrega- tion of conventional soil maps using database knowledge extraction to produce soil property maps, In: Digital Soil Assessments and Beyond: 5th Global Work- shop
Δ NA... Additionally, a crucial parameter for a coherent nonlinear effect such as THG is the spatial distribution of the phase. In line with previous reports [43], Fig. 2D shows
Edited by: F. C.: On the changing seasonal cycles and trends of ozone at Mace Head, Ireland, Atmos. S., Wang, T., and Giorgi, F.: Is ozone pollution affecting crop yields in
2013 made an assessment of impact of projected climate change on the hydrologic regime and climate extremes of the Congo River basin.. This specific river basin, despite its
Precipitation data were obtained from the GPCC project (Global Global Preciptiation Climatology.. Bahn et al.: Soil respiration at mean annual temperature predicts annual total
Differences in number of bumblebees, honeybees, mean total number of flowers and richness of species in flower per sampling date on the three seed mixtures in each year (ns,
There are several modi- fied live vaccines currently on the market for horses, including, but not limited to, vaccines for equine herpes, equine influ- enza, equine encephalitis,
relationship between NDVI time series and pearl millet aboveground biomass simulated by the crop 444.
