Control of Systemic Iron Homeostasis by the 3’ Iron-Responsive Element of Divalent Metal Transporter 1 in Mice
Texte intégral
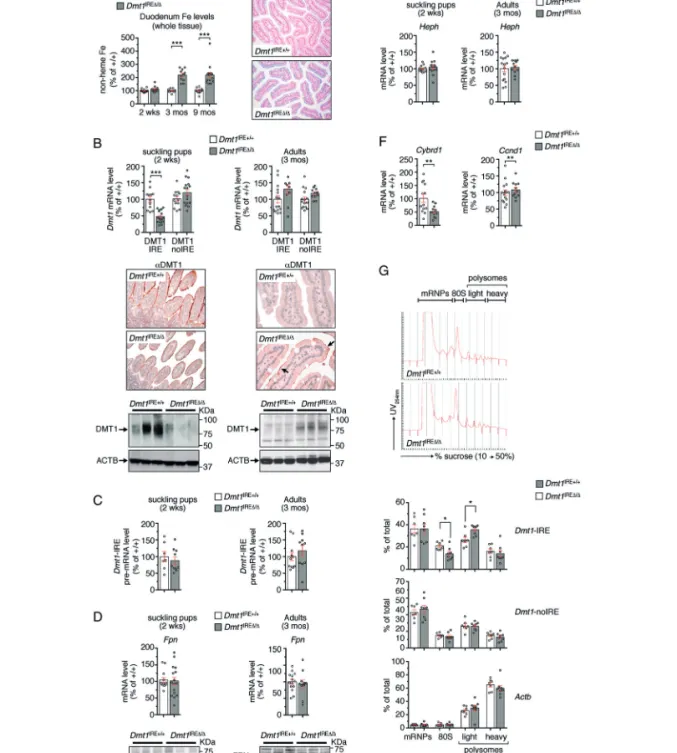
Figure

Documents relatifs
To overcome the intrinsic limitations of classical Genome-wide association studies (GWAS) involving univariate tests between statistically dependent markers, we have proposed in
Probably only this mixing causes initial absorption in ionic and covalent crystals, because in spectra of water soluted ions [4] without the effect of the crystal-
reported increased mitochondrial iron levels in HEK SFXN2 KO cells [9], an ICP-MS analysis did not show significantly modified cellular or mitochondrial iron levels in HEK SFXN1
In this context, by using a neonatal mouse model of cryptosporidiosis, we tried to decipher the intestinal consequences at adulthood of the neonatal infection
It must be emphasized that the scope of this note is confined solely to the fire hazard associated with paper- backed batts in enclosed ceiling spaces. The hazard associated with
The present study emphasized the effects of formulation process parameters on the characteristics and in vitro release behavior of CN-loaded microcapsules prepared
In the southern part of the Norwegian Sea the depth layers occu- pied by the species Calanus finmarchicus corresponds to the upper parts of Norwegian Sea deep water, a polar water
In the frame of this thesis, we have further investigated the molecular mechanisms involved in iron homeostasis in Pseudomonas aeruginosa (pathogen bacteria used as a model)
