Tectonic-driven climate change and the diversification of angiosperms
Texte intégral
Figure
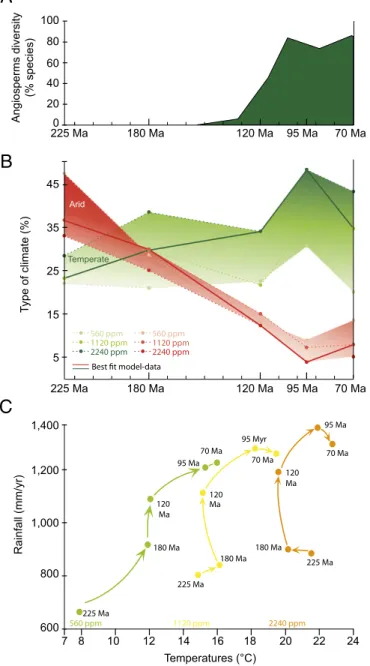
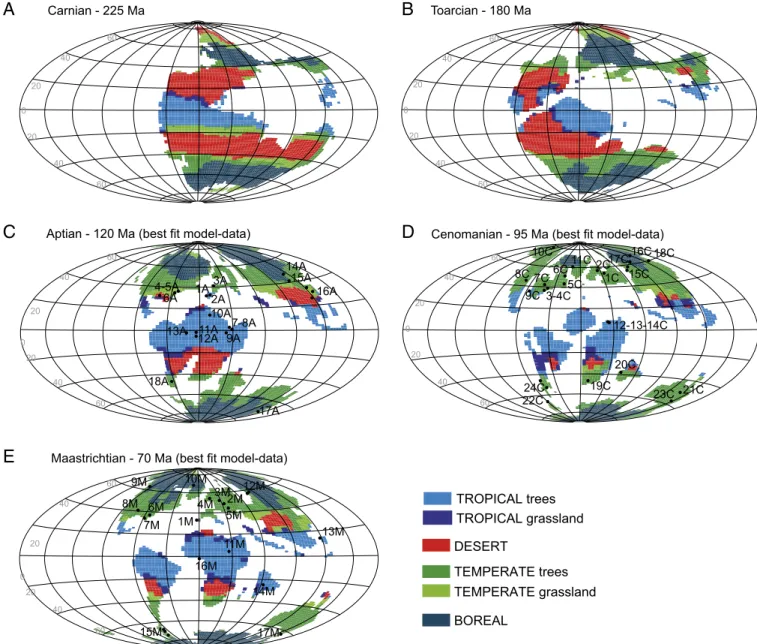
Documents relatifs
range expansions under climate change, the presence of moving climate envelopes can constrain the leading edge of the population in its expansion relative to the core of the
Therefore, the first-order evolution of the clay mineral associ- ation through the siliciclastic cover sequence can be interpreted as the result of (i) decrease of weathering
The increasing availability of high-resolution climatological and ecological reconstructions allows us to study the impact of past climate change on a human scale, one that is
This report on forests and climate change produced by the Central African Forest Commission COMIFAC with the support of its partners aims to update the international community and
Linder, 2017), evidence from vegetation and climate models, fossil sites, and dated molecular phylogenies favours the hypothesis of a rain forest band reconnecting east and west
➌ Change in the number of potentially favourable river basins for different species of European migratory fi sh. Blue indicates the number of river basins that will remain
In December 2010, Aéroports du Grand Ouest – the official name of the airport, and 85% owned by French transnational concession and construction conglomerate Vinci – signed a
Assessing the impact of climate variability and climate change on runoff in West Africa: The case of Senegal and Nakambe River basins.H. (wileyonlinelibrary.com)
