HAL Id: hal-02810535
https://hal.inrae.fr/hal-02810535
Submitted on 6 Jun 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of
sci-entific research documents, whether they are
pub-lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destinée au dépôt et à la diffusion de documents
scientifiques de niveau recherche, publiés ou non,
émanant des établissements d’enseignement et de
recherche français ou étrangers, des laboratoires
publics ou privés.
Evaluation in vitro du potentiel bioprotecteur des
champignons mycorhiziens à arbuscules contre le
flétrissement bactérien de la tomate
Patrice Crozilhac
To cite this version:
Patrice Crozilhac. Evaluation in vitro du potentiel bioprotecteur des champignons mycorhiziens à
arbuscules contre le flétrissement bactérien de la tomate. Sciences agricoles. 2013. �hal-02810535�
Année académique 2012-2013
Evaluation in vitro du potentiel bioprotecteur des
champignons mycorhiziens à arbuscules contre le
flétrissement bactérien de la tomate
-
Article de résumé
Présenté par Patrice Crozilhac
Promoteurs :
Prof. Stephane Declerck
1PhD. Student Katia Plouznikoff
1Ir. Marie Chave
2Lecteurs :
Prof. Claude Bragard (UCL/ELI/ELIM)
Prof. Xavier Draye (UCL/ELI/ISV/ELIA)
Article de résumé présenté pour les Crelan Awards
1,Earth and Life Institute, Mycology, Université catholique de Louvain, B-1348 Louvain-la-Neuve, Belgium
Article de résumé
Patrice Crozilhac - Année académique 2012-2013
1
Figure 1. Principaux mécanismes impliqués dans la bioprotection des plantes par les CMA (d’après
Patrice Crozilhac - Année académique 2012-2013
2
Le sol représente le premier réservoir de biodiversité1 mais cette biodiversité est encore peu valorisée dans les stratégies de gestion des agrosystèmes (Brussaard et al. 2007). Les microorganismes associés à la plante fournissent de multiples services écosystémiques parmi lesquels la bioprotection. Manipuler ces microorganismes pour accroître la santé et la productivité des agrosystèmes est un levier majeur pour réduire leur dépendance aux intrants chimiques (Ryan et al. 2009) et faciliter leur écologisation.
Pour accompagner la transition écologique, plusieurs plans, programmes et dispositifs ont émergé. En Belgique, par exemple, les Mesures Agro-environnementales (MAE) mises en œuvre au travers du Programme de Développement rural (PDR) visent notamment à réduire l’utilisation de pesticides tout en maintenant un niveau élevé de production agricole. La mise en œuvre par les agriculteurs de démarches d’intensification écologique conciliant compétitivité et respect de l’environnement est cependant encore sous exploitée (Guillou et al., 2013).
Un parasitisme tellurique exacerbé
Contrastées en termes d’enjeux (i. de préservation de la biodiversité, ii. de sécurité alimentaire, iii. de place de l’agriculture dans l’activité économique, iv. de sensibilisation des acteurs aux risques liés aux productions agricoles), les importantes régions agricoles Belges comme les Antilles Françaises sont des zones de forte production maraîchère où les solanacées (pomme de terre et tomate principalement) sont une spéculation majeure mais reste cependant fragilisée par un parasitisme tellurique exacerbé (bactéries, nématodes, champignons phytopathogènes) (Blancard, 2009).
Pour l’instant isolé aux régions tropicales et subtropicales, le flétrissement bactérien conduit à des pertes de récolte pouvant aller jusqu’à 100 %. Dans les régions tempérées, le contrôle des nématodes est également un véritable enjeu, suite aux restrictions d’emploi des nématicides chimiques (Wesemael et al., 2011). En France déjà près de 40% des exploitations légumières du Sud-Est sont touchés (Djian-Caporalino, 2010, 2012).
Une stratégie innovante de gestion des bioagresseurs
L’optimisation des rotations, en complémentarité de la gestion durable des résistances et du recours aux techniques culturales telles que la solarisation, les rotations de cultures non-hôtes ou assainissantes vis-à-vis des agents pathogènes majeurs rencontrés en cultures légumières plein sol, est au cœur des stratégies innovantes de gestion des bioagresseurs telluriques en zone tempérée comme en zone tropicale.
Certaines espèces, telles que l’oignon ou les crotalaires, au-delà de leurs propriétés directes sur les agents pathogènes, sont connues pour leur capacité à mobiliser et multiplier des microorganismes bioprotecteurs présents naturellement dans le sol, tels que les champignons mycorhiziens à arbuscules (CMA).
Les CMA sont des microorganismes fongiques de la division des Gloméromycètes, anciennement classés dans la famille des Zygomycètes (Schüßler et al., 2001). Ils forment des symbioses avec plus de 80% des espèces végétales (Smith and Read, 2008).
L’association symbiotique des CMA avec le système racinaire des plantes apporte une série de bénéfices à son hôte. Ces bénéfices multiples (multiservices) et bien référencés, sont qualifiés de services écosystémiques (Tableau 1).
Un des services écosystémiques majeur procuré par la symbiose mycorhizienne est la bioprotection des plantes hôtes contre certains pathogènes telluriques et aériens. Cette bioprotection est le résultat d’interactions complexes et parfois synergiques entre la plante, le pathogène, les communautés microbiennes de la rhizosphère et le CMA (Figure 1)
Parmi les nombreuses espèces d’intérêt agronomique qui bénéficient de la symbiose mycorhizienne, la pomme de terre et la tomate font parties de celles qui maximise l’effet bioprotecteur des CMA (Gallou et al, 2010; Gianinazzi et al, 2010).
11 gramme de sol rhizosphérique peut contenir jusqu’à 1011 microorganismes de plus de 30 000 espèces différentes (Berendsen et al. 2012).
Article de résumé
Patrice Crozilhac - Année académique 2012-2013
3
Les CMA, induisent un biocontrôle caractérisé par la réduction des symptômes des maladies provoquées principalement par les champignons, les nématodes et les bactéries phytopathogènes du sol.
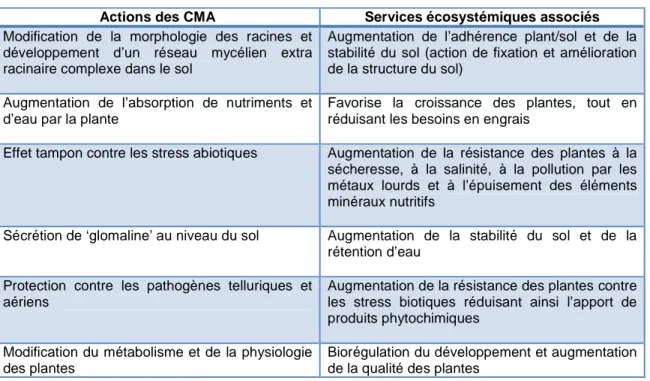
Tableau 1 : Services écosystémiques rendus par les CMA, inspiré de Smith & Read (2008)
Actions des CMA Services écosystémiques associés
Modification de la morphologie des racines et développement d’un réseau mycélien extra racinaire complexe dans le sol
Augmentation de l’adhérence plant/sol et de la stabilité du sol (action de fixation et amélioration de la structure du sol)
Augmentation de l’absorption de nutriments et d’eau par la plante
Favorise la croissance des plantes, tout en réduisant les besoins en engrais
Effet tampon contre les stress abiotiques Augmentation de la résistance des plantes à la sécheresse, à la salinité, à la pollution par les métaux lourds et à l’épuisement des éléments minéraux nutritifs
Sécrétion de ‘glomaline’ au niveau du sol Augmentation de la stabilité du sol et de la rétention d’eau
Protection contre les pathogènes telluriques et aériens
Augmentation de la résistance des plantes contre les stress biotiques réduisant ainsi l’apport de produits phytochimiques
Modification du métabolisme et de la physiologie des plantes
Biorégulation du développement et augmentation de la qualité des plantes
La tomate : Une ‘cash crop’ modèle
Au-delà de son importance économique, du bénéfice fourni par les CMA et de son statut de plante modèle, la tomate est une culture qui présente de nombreux avantages pour l’étude d’une telle stratégie de lutte, transposable à d’autres cultures : cycle court, diversité très large de systèmes de production (de l’agroforesterie à l’hydroponie), retour sur investissement potentiellement rapide pour l’agriculteur.
Des processus à décrypter pour optimiser la mise en œuvre des plantes mycorhizotrophes et assainissantes pour la bioprotection in situ.
L’objectif de ce travail était de déterminer comment associer des plantes potentiellement mycorhizotrophes et biorégulatrices à la tomate pour la bioprotéger contre le flétrissement bactérien. La bactérie tellurique R. solanacearum, responsable du flétrissement bactérien de la tomate, constitue une contrainte biotique majeure dans les systèmes de cultures maraîchères en Martinique comme dans toute la ceinture tropicale et subtropicale humide.
Parmi les espèces candidates, celles du genre Allium spp et Crotalaria spp ont été étudiées comme plantes de services pour leurs potentiels biorégulateur et mycorhizotrophe.
L’évaluation en microcosme in vitro des processus de bioprotections impliqués est possible grâce aux systèmes mis en place au sein du laboratoire de mycologie de l’UCL par Voets (2009). Ces systèmes permettent l’étude des interactions des plantes mycorhizées avec des agents pathogènes en condition monoxénique et de façon non destructive.
La stratégie expérimentale qui a été mise en œuvre a consisté à évaluer dans des systèmes de culture in vitro associant une plantule de tomate et un CMA, le potentiel bioprotecteur des CMA contre
Patrice Crozilhac - Année académique 2012-2013
4
Cette expérience principale a reposée sur deux expériences préliminaires qui ont permis d’évaluer d’une part le potentiel de Crotalaria spectabilis et Allium fistulosum comme plantes-donneuses de mycorhizes pour la tomate et d’évaluer d’autre part la dynamique de colonisation des plants de tomate par un CMA. L’incidence du flétrissement au cours du temps a été mesurée selon deux échelles différentes (le pourcentage de plantes flétries (PWP) et l’échelle de He (DI)). Ces expériences ont été complétées par une expérience exploratoire, visant à étudier le comportement in vitro de C. spectabilis et A. fistulosum mycorhizés en présence de R. solanacearum.
L’analyse in vitro, de la dynamique de colonisation des plants de tomates par un CMA, a permis de mettre en évidence une colonisation maximum des racines (20 ± 3%) par le CMA en 13 jours.
Les résultats obtenus in vitro, montre que la pré-mycorhization des plants de tomates par le CMA
Rhyzophagus irregularis MUCL 41833 diminue l’incidence du flétrissement causé par R. solanacearum.
En effet, les mesures d’AUDPC, qui intègrent la progression du flétrissement pour chaque traitement, ont permis d’évaluer la sévérité de la maladie sous 18 jours.
Une diminution de la sévérité de la maladie de 20% (± 0.4%) et de 21% (± 0.5%), selon respectivement l’indice de flétrissement (DI) et le pourcentage de plantes flétries (PWP) a ainsi été observée suite à une phase de pré-mycorhization des plants de tomates.
Ces observations ont permis de confirmer l’effet bioprotecteur des CMA et de mettre en évidence, pour la première fois, la capacité de biocontrôle du R. irregularis MUCL 41833, chez des plants de tomates infestés par R. solanacearum.
Ces résultats s’expliquent par différents mécanismes qui peuvent être synergiques. Ainsi les hypothèses de compétition pour les sites de colonisations, d’induction de mécanismes de défense et de modifications de la composition biochimique des exsudats, ont été discutés, comme facteurs de bioprotection, induits par les CMA.
Une étape indispensable à la mise en place de stratégies de gestion agroécologique.
Ces recherches s’insèrent comme une piste originale et porteuse quant à la manière d’aborder l’insertion des plantes de services mycorhizotrophes et à propriétés allélopathiques dans une rotation culturale. Les résultats obtenus permettent de prétendre à une maitrise plus fine du potentiel bioprotecteur des CMA contre le flétrissement bactérien de la tomate et ouvre ainsi un voie générique d’utilisation des CMA comme agents de biocontrôles contre les pathogènes telluriques et aériens en cultures maraichères et horticoles.
Références bibliographiques
Berendsen R L, Pieterse CMJ and Bakker PAHM. (2012). The rhizosphere microbiome and plant
health. Trends Plant Sci. 17: 478-486.
Blancard, D. (2009). Les maladies de la tomate: identifier, connaître, maîtriser. Quae éditions. pp. 679 Brussaard L., de Ruiter P. C., Brown G. G., (2007). Soil biodiversity for agricultural sustainaibility.
Agriculture, Ecosystems and Environment 121 : 233-244
Djian-Caporalino C., (2010). Nématodes à galles, des ravageurs de plus en plus préoccupants :
résultats de trois ans d’enquête dans quinze régions françaises. Phytoma –La défense des végétaux, 638: 43-49
Djian-Caporalino C., (2012). Root-knot nematodes (Meloidogyne spp.), a growing problem in French
vegetable crops. EPPO Bulletin 42 (1): 127-137.
Gallou, A., De Jaeger, N., Cranenbrouck, S., & Declerck, S. (2010). Fast track in vitro mycorrhization
of potato plantlets allow studies on gene expression dynamics. Mycorrhiza, 20(3), pp. 201-207.
Gianinazzi S., Golotte A., Binet M-N., Van Tuinen D., Redecker D., Wipf D. (2010). Agroecology: the
key role of mycorrhizas in ecosystem services. Mycorrhiza.
Guillou M., Guyomard H., Huyghe C., Peyraud J-L. (2013). Le projet agro-écologique: vers des
agricultures doublement performantes pour concilier compétitivité et respect de l’environnement. Propositions pour le ministre de l’Agriculture, de l’agro-alimentaire et de la Forêt remis le 11 Juin 2013. 163 p.
Ryan P. R., Dessaux Y., Thomashow L.S., Weller, D. M. (2009). Rhizosphere engineering and
Article de résumé
Patrice Crozilhac - Année académique 2012-2013
5
Schüßler, A., Schwarzott, D., & Walker, C. (2001). A new fungal phylum the Glomeromycota:
phylogeny and evolution. Mycological Research 105, pp. 1413-1421.
Smith S and Read J (2008) Mycorrhizal symbiosis (Ed. Hardcover), 800 p
Voets L., Providencia I.E., Fernandez K., Ijdo M., Cranenbrouck S., Declerck S. 2009. Extraradical
mycelium network of arbuscular mycorrhizal fungi allows fast colonization of seelings under in vitro conditions. Mycorrhiza.
Wesemael W.M.L., Viaene N., Moens M., (2011). Root-knot nematodes (Meloidogyne spp.) in