Classical biological control: exploiting enemy escape to manage plant invasions
16
0
0
Texte intégral
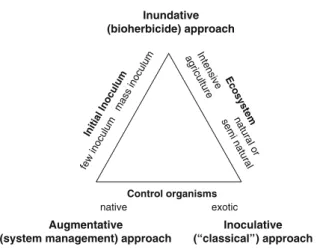
Figure

Documents relatifs