Quantifying the Effects of 16p11.2 Copy Number Variants on Brain Structure: A Multisite Genetic-First Study
Texte intégral
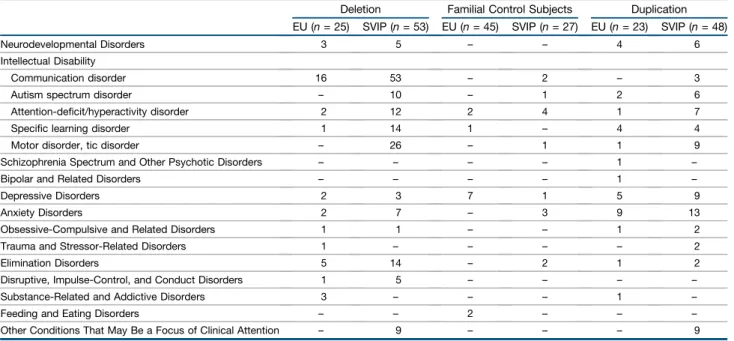
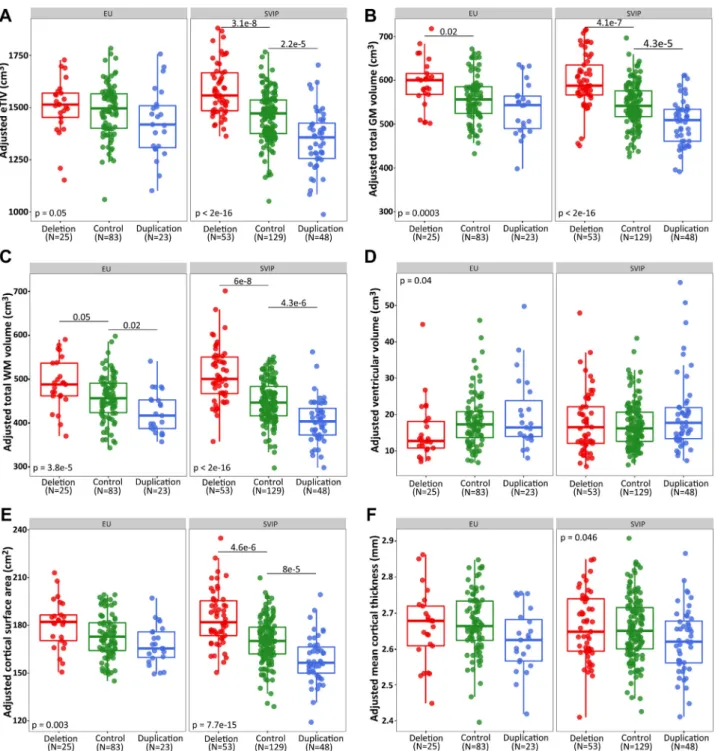
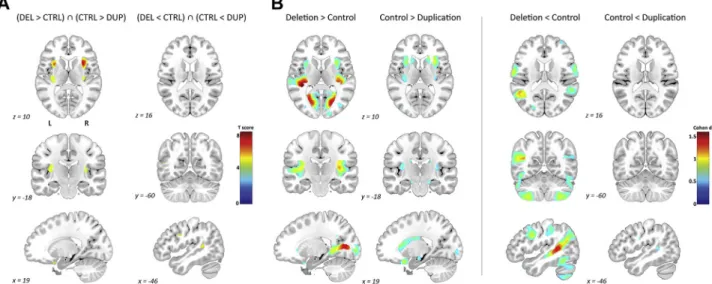
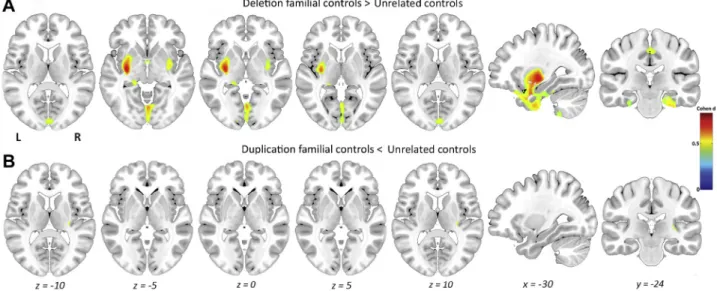
Figure
Documents relatifs
Thrombosis prevention trial: randomized trial of low-intensity oral anticoagulation with warfa- rin and low-dose aspirin in the primary prevention of ischemic heart disease in men
Conformément à l’arrêté du 31 décembre 2004 modifié, PAPRICA effectue les traitements suivants pour produire ce type de fichier : inscription des versions,
Tammy souhaitait avoir une discussion sérieuse depuis lundi
neigh-neigh
Une formation courte est également dispensée aux chefs de clinique (tous les 6 mois car le turnover est important). Le Dr Esterle et le sociologue Mathieu-Fritz ont étudié dans
This is an integrated initiative of four United Nations agencies (United Nations Children’s Fund, United Nations Educational, Scientific and Cultural Organization, United
Jean-yves rEyMonD, cadre supérieur de santé, direction de la qualité-gestion des risques, aP-hM Qualité de vie en HAD et témoignage de familles thibaut lEFEBvrE, interne et Dr
The generalisation of employer-sponsored Complementary Health Insurance may have two effects: on the one hand, an end to residual situations in which there is an absence of
