Highly aligned lamellar lipid domains induced by macroscopic confinement
Texte intégral
Figure
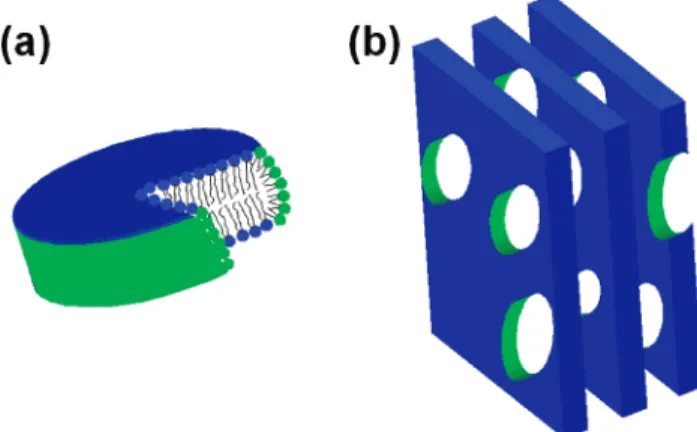
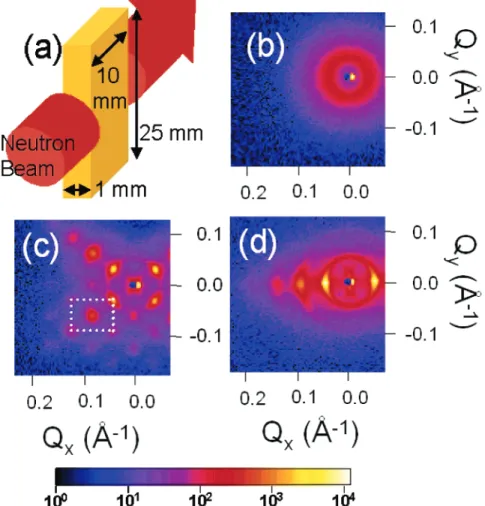
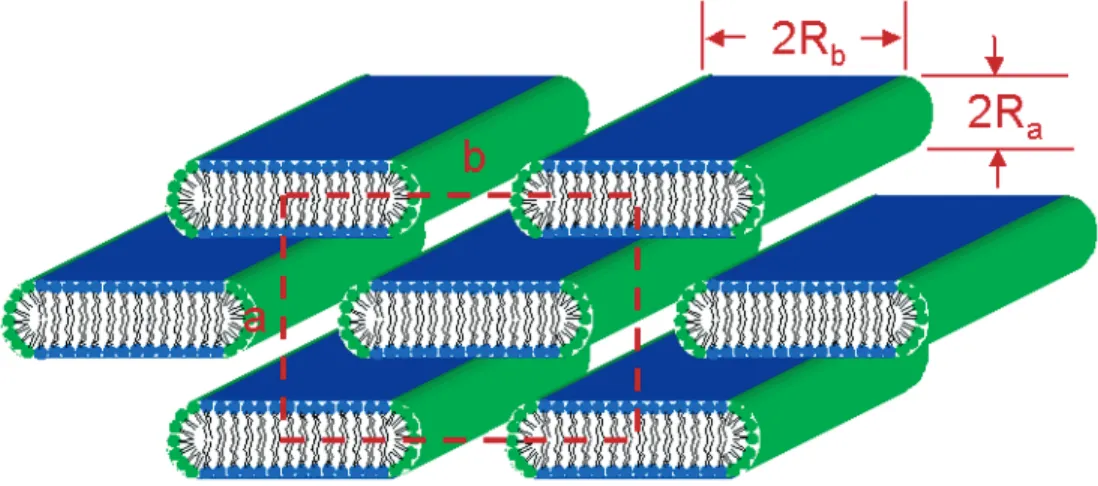


Documents relatifs
Composition Au cours des années 2006 et 2007, les personnes suivantes ont été membres du Conseil de bibliothèque : l’administrateur en charge de la politique institutionnelle
Ectopic Bmp2 expression in the ventricles increased pSmad1/5 levels, resulting in transcriptional activation of Notch ligands, expansion of Notch pathway activity, and
In the large /I case (g uc) three different phases already exist at zero shear rate : the homogeneous disordered phase at high temperatures, the hexagonal phase
In order to decide whether non-stationary tilt inclusions may exist for values of p different from p *, as well as to check whether these can give Rse to dynamical selection, we
This Apollonian weak metric has been studied in the paper [5], in relation with a weak metric introduced by Thurston on the Teichm¨ uller space of a hyperbolic surface [17] and it
Equation (12) has all the features required to describe a second-order lamellar-to-cubic phase transition. the average principal curvatures of the bilayer. However,
R6sumi: Etant donnd le caractbre bidimensionnel des materiaux lamellaires, les spectres d'absorption des rayons X sont trbs sensibles aux effets de polarisation qui
A linear stability analysis shows that unstable modulations develop at low and intermediate pressures, following an instability mechanism due to an energy transport effect —
